A Complete End-to-End Workflow for untargeted LC-MS/MS Metabolomics Data Analysis in R
Philippine Louail, Anna Tagliaferri, Vinicius Verri Hernandes, Daniel M. S. Silva, Johannes Rainer
2023-09-07
Source:vignettes/end-to-end-untargeted-metabolomics.Rmd
end-to-end-untargeted-metabolomics.RmdAbstract
Metabolomics provides a real-time view of the metabolic state of examined samples, with mass spectrometry serving as a key tool in deciphering intricate differences in metabolomes due to specific factors. In the context of metabolomic investigations, untargeted liquid chromatography with tandem mass spectrometry (LC-MS/MS) emerges as a powerful approach thanks to its versatility and resolution. This paper focuses on a dataset aimed at identifying differences in plasma metabolite levels between individuals suffering from a cardiovascular disease and healthy controls.
Despite the potential insights offered by untargeted LC-MS/MS data, a significant challenge in the field lies in the generation of reproducible and scalable analysis workflows. This struggle is due to the aforementioned high versatility of the technique, which results in difficulty in having a one-size-fits-all workflow and software that will adapt to all experimental setups. The power of R-based analysis workflows lies in their high customizability and adaptability to specific instrumental and experimental setups; however, while various specialized packages exist for individual analysis steps, their seamless integration and application to large cohort datasets remain elusive. Addressing this gap, we present an innovative R workflow that leverages xcms, and packages of the RforMassSpectrometry environment to encompass all aspects of pre-processing and downstream analyses for LC-MS/MS datasets in a reproducible manner and allow an easy customization to generate data-set specific workflows. Our workflow seamlessly integrates with Bioconductor packages, offering adaptability to diverse study designs and analysis requirements.
Keyword
LC-MS/MS, reproducibility, workflow, xcms, R, normalization, feature identification, Bioconductor,…
Introduction
Liquid chromatography coupled to tandem mass spectrometry (LC-MS/MS) is a powerful tool for metabolomics investigations, providing a comprehensive view of the metabolome. It enables the identification of a large number of metabolites and their relative abundance in biological samples. Liquid Chromatography (LC) is a separation technique which relies on the different interactions of the analytes towards a chromatographic column - the stationary phase - and the eluent of the analysis - the mobile phase. The stronger the affinity of an analyte to the stationary phase - dictated by polarity, size, charges and other parameters - the longer it will take for the compound to leave the column and be detected by our coupled technique - the Mass Spectrometer.
Mass Spectrometry allows to identify and quantify ions based on their mass-to-charge (m/z) ratio. Its high selectivity relies not only on the capability to separate compounds with very small variations in mass, but also on the capacity to promote fragmentation. An ion with an initial m/z (the parent ion) can be broken into characteristic fragments (daughter ions), which would then help in the structure elucidation and identification of that specific compound (Theodoridis et al. 2012).
Therefore, LC-MS/MS data are usually tridimensional datasets containing the retention time of the compounds after separation by LC, the detected m/z of all the compounds at a given time, and the intensity of each of such signals. Furthermore, each MS signal can have two different levels, corresponding to the signal of the parent ion (called MS1) and the signals from their corresponding fragments (denominanted MS2).
The high sensitivity and specificity of LC-MS/MS make it an indispensable tool for biomarker discovery and elucidating metabolic pathways. The untargeted approach is particularly useful for hypothesis-free investigations, allowing for the detection of unexpected metabolites and pathways. However, the analysis of LC-MS/MS data is complex and requires a series of preprocessing steps to extract meaningful information from the raw data. The main challenges include dealing with the lack of ground truth data, the high dimensionality of the data, and the presence of noise and artifacts (Gika, Wilson, and Theodoridis 2014). Moreover, due to different instrumental setups and protocols the definition of a single one-fits-all workflow is impossible. Finally, while specialized software packages exist for each individual step during an analysis, their seamless integration remains elusive.
Here we present a complete analysis workflow for untargeted LC-MS/MS data using R and Bioconductor packages, in particular those from the RforMassSpectrometry package ecosystem. The later is an initiative initiative that aims to implement an expandable, flexible infrastructure for the analysis of MS data, providing also a comprehensive toolbox of functions to build customized analysis workflows. We demonstrate how the various algorithms can be adapted to the particular data set and how various R packages can be seamlessly integrated to ensure efficient and reproducible processing. The present workflow covers all steps for LC-MS/MS data analysis, from preprocessing, data normalization, differential abundance analysis to the annotation of the significant features i.e., collections of signals that have the same retention time and mass-to-charge ratios pertaining to the same ions. Various options for visualizations as well as quality assessment are presented for all analysis steps.
Data description
In this workflow two datasets are utilized, an LC-MS-based (MS1 level only) untargeted metabolomics data set to quantify small polar metabolites in human plasma samples and an additional LC-MS/MS data set of selected samples from the former study for the identification/annotation of its significant features. The samples used here were randomly selected from a larger study for the identification of metabolites with differences in abundances between individuals suffering from a cardiovascular disease (CVD) and healthy controls (CTR).The subset analyzed here comprises data for three CVD and three CTR as well as four quality control (QC) samples. The QC samples represent a pool of serum samples from a large cohort and were repeatedly measured throughout the experiment to monitor stability of the signal.
The data and metadata for this workflow are accessible on the MetaboLight database under the ID: MTBLS8735.
The detailed materials and method used for the analysis of the samples can also be found on the metabolight database. As it is especially pertinent for the analysis and the chosen parameters, we want to highlight that the samples were analyzed using ultra-high-performance liquid chromatography (UHPLC) coupled to a Q-TOF mass spectrometer (TripleTOF 5600+). The chromatographic separation was based on hydrophilic interaction liquid chromatography (HILIC).
Workflow description
The present workflow describes all steps for the analysis of an LC-MS/MS experiment, which includes the preprocessing of the raw data to generate the abundance matrix for the features in the various samples, followed by data normalization, differential abundance analysis and finally the annotation of features to metabolites. Note that also alternative analysis options and R packages could be used for different steps and some examples are mentioned throughout the workflow. [jo: I’ll include some of these maybe later. It would be key to justify why this workflow is comprehensive]
Our workflow is therefore based on the following dependencies:
## General bioconductor package
library(Biobase)
## Data Import and handling
library(readxl)
library(MsExperiment)
library(MsIO)
library(MsBackendMetaboLights)
library(SummarizedExperiment)
## Preprocessing of LC-MS data
library(xcms)
library(Spectra)
library(MetaboCoreUtils)
## Statistical analysis
library(limma) # Differential abundance
library(matrixStats) # Summaries over matrices
## Visualisation
library(pander)
library(RColorBrewer)
library(pheatmap)
library(vioplot)
library(ggfortify) # Plot PCA
library(gridExtra) # To arrange multiple ggplots into single plots
## Annotation
library(AnnotationHub) # Annotation resources
library(CompoundDb) # Access small compound annotation data.
library(MetaboAnnotation) # Functionality for metabolite annotation.Data import
Note that different equipment will generate various file extensions, so a conversion step might be needed beforehand, though it does not apply to this dataset. The Spectra package supports a variety of ways to store and retrieve MS data, including mzML, mzXML, CDF files, simple flat files, or database systems. If necessary, several tools, such as ProteoWizard’s MSConvert, can be used to convert files to the .mzML format (Chambers et al. 2012).
Below we will show how to extract our dataset from the MetaboLigths
database and load it as an MsExperiment object. For more
information on how to load your data from the MetaboLights database,
refer to the vignette.
For other type of data loading, check out this link:
param <- MetaboLightsParam(mtblsId = "MTBLS8735",
assayName = paste0("a_MTBLS8735_LC-MS_positive_",
"hilic_metabolite_profiling.txt"),
filePattern = ".mzML")
data <- readMsObject(MsExperiment(),
param,
keepOntology = FALSE,
keepProtocol = FALSE,
simplify = TRUE)We next configure the parallel processing setup. Most functions from the xcms package allow per-sample parallel processing, which can improve the performance of the analysis, especially for large data sets. xcms and all packages from the RforMassSpectrometry package ecosystem use the parallel processing setup configured through the BiocParallel Bioconductor package. With the code below we use a fork-based parallel processing on unix system, and a socket-based parallel processing on the Windows operating system.
#' Set up parallel processing using 2 cores
if (.Platform$OS.type == "unix") {
register(MulticoreParam(2))
} else{
register(SnowParam(2))
}Data organisation
The experimental data is now represented by a
MsExperiment object from the MsExperiment
package. The MsExperiment object is a container for
metadata and spectral data that provides and manages also the linkage
between samples and spectra.
data## Object of class MsExperiment
## Spectra: MS1 (17210)
## Experiment data: 10 sample(s)
## Sample data links:
## - spectra: 10 sample(s) to 17210 element(s).Below we provide a brief overview of the data structure and content.
The sampleData() function extracts sample information from
the object. We next extract this data and use the pander
package to render and show this information in Table 1 below. Throughout
the document we use the R pipe operator (|>) to avoid
nested function calls and hence improve code readability.
| Derived_Spectral_Data_File | Characteristics.Sample.type. |
|---|---|
| FILES/MS_QC_POOL_1_POS.mzML | pool |
| FILES/MS_A_POS.mzML | experimental sample |
| FILES/MS_B_POS.mzML | experimental sample |
| FILES/MS_QC_POOL_2_POS.mzML | pool |
| FILES/MS_C_POS.mzML | experimental sample |
| FILES/MS_D_POS.mzML | experimental sample |
| FILES/MS_QC_POOL_3_POS.mzML | pool |
| FILES/MS_E_POS.mzML | experimental sample |
| FILES/MS_F_POS.mzML | experimental sample |
| FILES/MS_QC_POOL_4_POS.mzML | pool |
| Factor.Value.Phenotype. | Sample.Name | Factor.Value.Age. |
|---|---|---|
| POOL | NA | |
| CVD | A | 53 |
| CTR | B | 30 |
| POOL | NA | |
| CTR | C | 66 |
| CVD | D | 36 |
| POOL | NA | |
| CTR | E | 66 |
| CVD | F | 44 |
| POOL | NA |
| derived_spectra_data_file | phenotype | sample_name | age |
|---|---|---|---|
| FILES/MS_QC_POOL_1_POS.mzML | QC | POOL1 | NA |
| FILES/MS_A_POS.mzML | CVD | A | 53 |
| FILES/MS_B_POS.mzML | CTR | B | 30 |
| FILES/MS_QC_POOL_2_POS.mzML | QC | POOL2 | NA |
| FILES/MS_C_POS.mzML | CTR | C | 66 |
| FILES/MS_D_POS.mzML | CVD | D | 36 |
| FILES/MS_QC_POOL_3_POS.mzML | QC | POOL3 | NA |
| FILES/MS_E_POS.mzML | CTR | E | 66 |
| FILES/MS_F_POS.mzML | CVD | F | 44 |
| FILES/MS_QC_POOL_4_POS.mzML | QC | POOL4 | NA |
| injection_index |
|---|
| 1 |
| 2 |
| 3 |
| 4 |
| 5 |
| 6 |
| 7 |
| 8 |
| 9 |
| 10 |
There are 11 samples in this data set. Below are abbreviations essential for proper interpretation of this metadata information:
- injection_index: An index representing the order (position) in which an individual sample was measured (injected) within the LC-MS measurement run of the experiment.
-
phenotype: The sample groups of the experiment:
-
"QC": Quality control sample (pool of serum samples from an external, large cohort). -
"CVD": Sample from an individual with a cardiovascular disease. -
"CTR": Sample from a presumably healthy control.
-
- sample_name: An arbitrary name/identifier of the sample.
- age: The (rounded) age of the individuals.
We will define colors for each of the sample groups based on their sample group using the RColorBrewer package:
The MS data of this experiment is stored as a Spectra
object (from the Spectra
Bioconductor package) within the MsExperiment object and
can be accessed using spectra() function. Each element in
this object is a spectrum - they are organised linearly and are all
combined in the same Spectra object one after the other
(ordered by retention time and samples).
Below we access the dataset’s Spectra object which will
summarize its available information and provide, among other things, the
total number of spectra of the data set.
#' Access Spectra Object
spectra(data)## MSn data (Spectra) with 17210 spectra in a MsBackendMetaboLights backend:
## msLevel rtime scanIndex
## <integer> <numeric> <integer>
## 1 1 0.274 1
## 2 1 0.553 2
## 3 1 0.832 3
## 4 1 1.111 4
## 5 1 1.390 5
## ... ... ... ...
## 17206 1 479.052 1717
## 17207 1 479.331 1718
## 17208 1 479.610 1719
## 17209 1 479.889 1720
## 17210 1 480.168 1721
## ... 36 more variables/columns.
##
## file(s):
## MS_QC_POOL_1_POS.mzML
## MS_A_POS.mzML
## MS_B_POS.mzML
## ... 7 more filesWe can also summarize the number of spectra and their respective MS
level (extracted with the msLevel() function). The
fromFile() function returns for each spectrum the index of
its sample (data file) and can thus be used to split the information (MS
level in this case) by sample to further summarize using the base R
table() function and combine the result into a matrix. Note
that this is the number of spectra acquired at each run, and not the
number of spectral features in each sample.
#' Count the number of spectra with a specific MS level per file.
spectra(data) |>
msLevel() |>
split(fromFile(data)) |>
lapply(table) |>
do.call(what = cbind)## 1 2 3 4 5 6 7 8 9 10
## 1 1721 1721 1721 1721 1721 1721 1721 1721 1721 1721The present data set thus contains only MS1 data, which is ideal for quantification of the signal. A second (LC-MS/MS) data set also with fragment (MS2) spectra of the same samples will be used later on in the workflow.
Note that users should not restrict themselves to data evaluation examples shown here or in other tutorials. The Spectra package enables user-friendly access to the full MS data and its functionality should be extensively used to explore, visualize and summarize the data.
As another example, we below determine the retention time range for the entire data set.
## [1] 0.273 480.169Data obtained from LC-MS experiments are typically analyzed along the retention time axis, while MS data is organized by spectrum, orthogonal to the retention time axis.
Data visualization and general quality assessment
Effective visualization is paramount for inspecting and assessing the quality of MS data. For a general overview of our LC-MS data, we can:
- Combine all mass peaks from all (MS1) spectra of a sample into a single spectrum in which each mass peak then represents the maximum signal of all mass peaks with a similar m/z. This spectrum might then be called Base Peak Spectrum (BPS), providing information on the most abundant ions of a sample.
- Aggregate mass peak intensities for each spectrum, resulting in the Base Peak Chromatogram (BPC). The BPC shows the highest measured intensity for each distinct retention time (hence spectrum) and is thus orthogonal to the BPS.
- Sum the mass peak intensities for each spectrum to create a Total Ion Chromatogram (TIC).
- Compare the BPS of all samples in an experiment to evaluate similarity of their ion content.
- Compare the BPC of all samples in an experiment to identify samples with similar or dissimilar chromatographic signal.
In addition to such general data evaluation and visualization, it is also crucial to investigate specific signal of e.g. internal standards or compounds/ions known to be present in the samples. By providing a reliable reference, internal standards help achieve consistent and accurate analytical results.
Spectra Data Visualization: BPS
The BPS collapses data in the retention time dimension and reveals the most prevalent ions present in each of the samples, creation of such BPS is however not straightforward. Mass peaks, even if representing signals from the same ion, will never have identical m/z values in consecutive spectra due to the measurement error/resolution of the instrument.
Below we use the combineSpectra function to combine all
spectra from one file (defined using parameter
f = fromFile(data)) into a single spectrum. All mass peaks
with a difference in m/z value smaller than 3 parts-per-million
(ppm) are combined into one mass peak, with an intensity representing
the maximum of all such grouped mass peaks. To reduce memory
requirement, we in addition first bin each spectrum combining
all mass peaks within a spectrum, aggregating mass peaks into bins with
0.01 m/z width. In case of large datasets, it is also
recommended to set the processingChunkSize() parameter of
the MsExperiment object to a finite value (default is
Inf) causing the data to be processed (and loaded into
memory) in chunks of processingChunkSize() spectra. This
can reduce memory demand and speed up the process.
#' Setting the chunksize
chunksize <- 1000
processingChunkSize(spectra(data)) <- chunksizeWe can now generate BPS for each sample and plot()
them.
Here, there is observable overlap in ion content between the files, particularly around 300 m/z and 700 m/z. There are however also differences between sets of samples. In particular, BPS 1, 4, 7 and 10 (counting row-wise from left to right) seem different than the others. In fact, these four BPS are from QC samples, and the remaining six from the study samples. The observed differences might be explained by the fact that the QC samples are pools of serum samples from a different cohort, while the study samples represent plasma samples, from a different sample collection.
Next to the visual inspection above, we can also calculate and
express the similarity between the BPS with a heatmap. Below we use the
compareSpectra() function to calculate pairwise
similarities between all BPS and use then the pheatmap()
function from the pheatmap package to cluster and visualize
this result.
We get a first glance at how our different samples distribute in terms of similarity. The heatmap confirms the observations made with the BPS, showing distinct clusters for the QCs and the study samples, owing to the different matrices and sample collections.
It is also strongly recommended to delve deeper into the data by exploring it in more detail. This can be accomplished by carefully assessing our data and extracting spectra or regions of interest for further examination. In the next chunk, we will look at how to extract information for a specific spectrum from distinct samples.
#' Accessing a single spectrum - comparing with QC
par(mfrow = c(1,2), mar = c(2, 2, 2, 2))
spec1 <- spectra(data[1])[125]
spec2 <- spectra(data[3])[125]
plotSpectra(spec1, main = "QC sample")
plotSpectra(spec2, main = "CTR sample")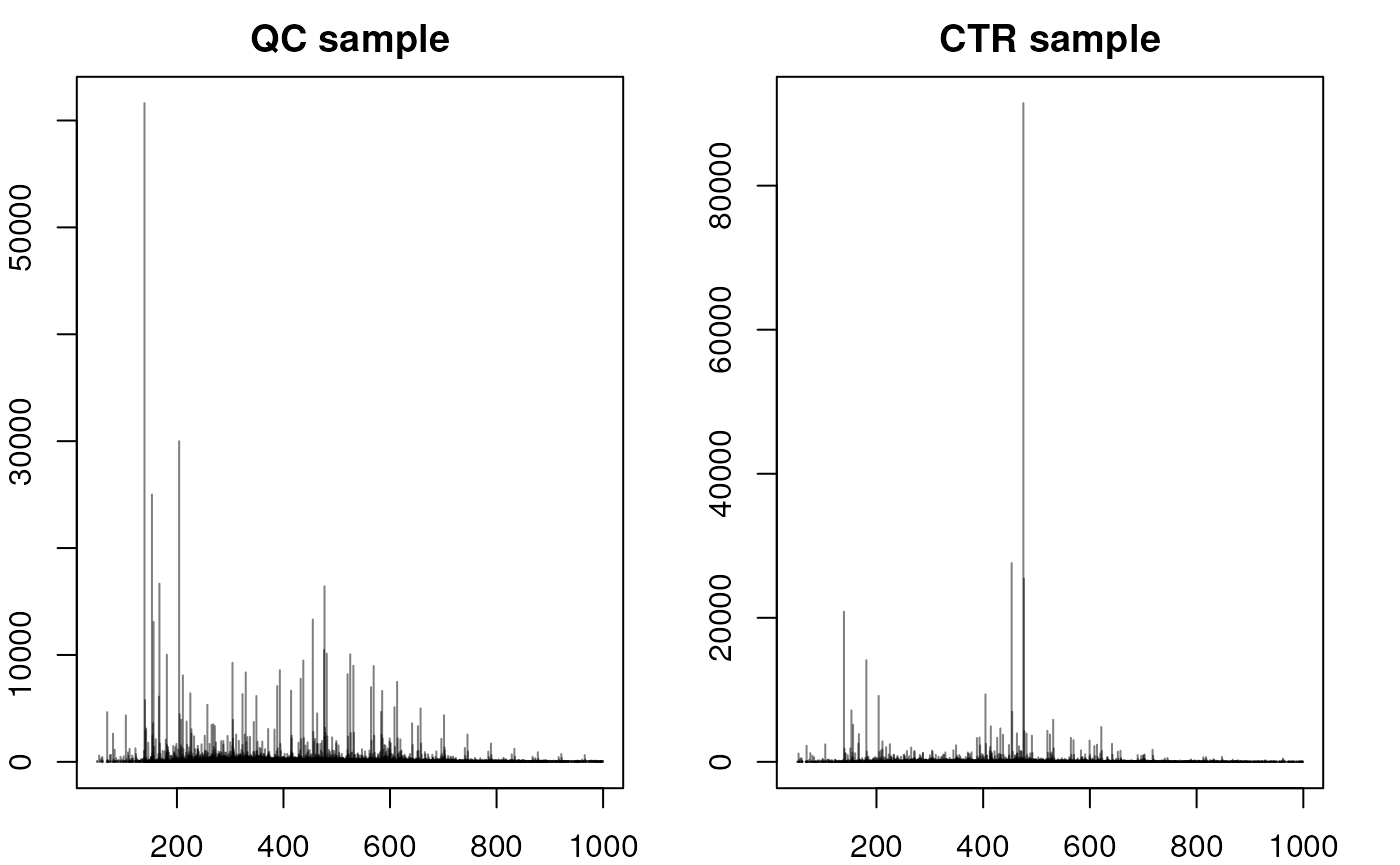
The significant dissimilarities in peak distribution and intensity confirm the difference in composition between QCs and study samples. We next compare a full MS1 spectrum from a CVD and a CTR sample.
#' Accessing a single spectrum - comparing CVD and CTR
par(mfrow = c(1,2), mar = c(2, 2, 2, 2))
spec1 <- spectra(data[2])[125]
spec2 <- spectra(data[3])[125]
plotSpectra(spec1, main = "CVD sample")
plotSpectra(spec2, main = "CTR sample")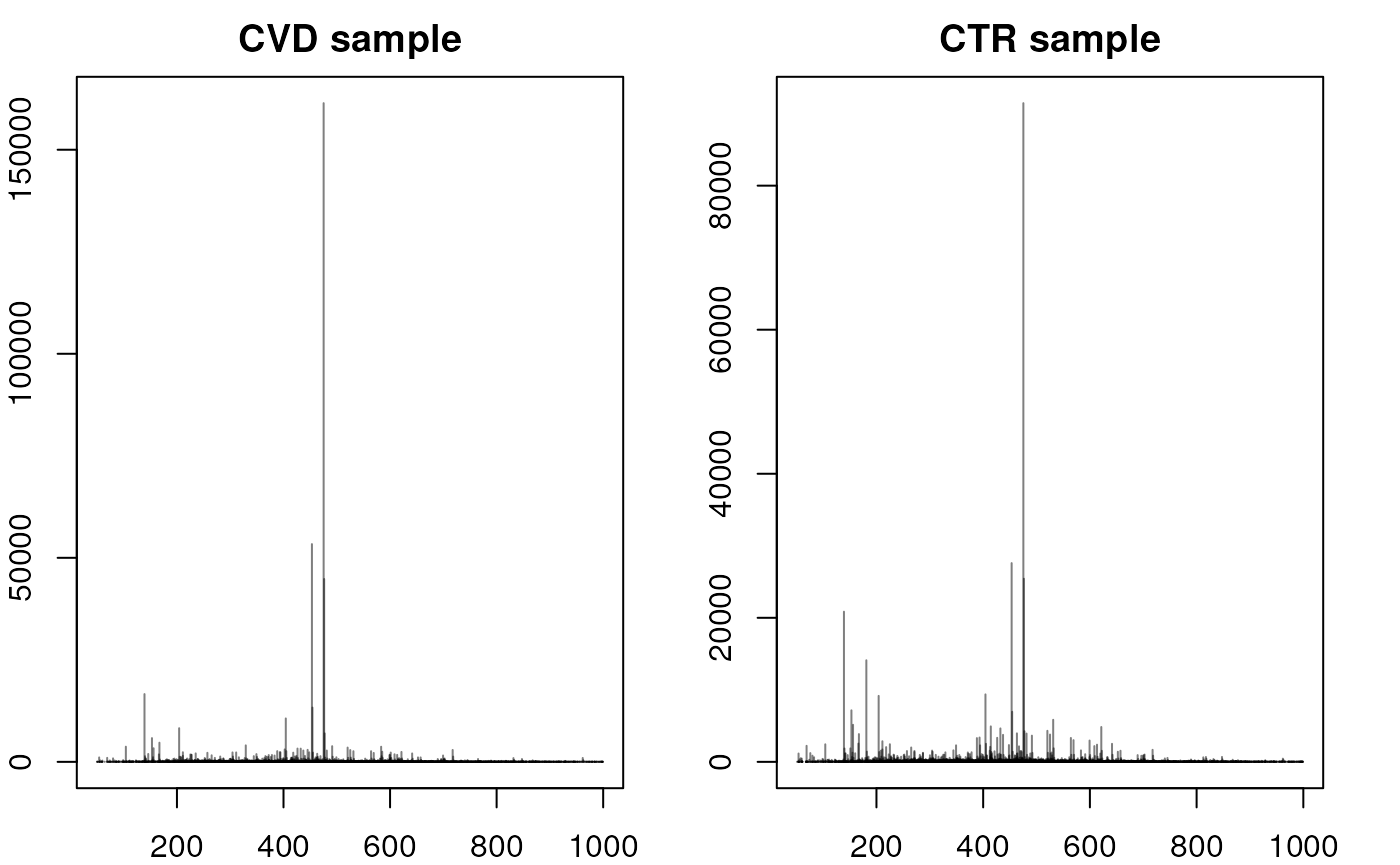
Above, we can observe that the spectra between CVD and CTR samples are not entirely similar, but they do exhibit similar main peaks between 200 and 600 m/z with a general higher intensity in control samples. However the peak distribution (or at least intensity) seems to vary the most between an m/z of 10 to 210 and after an m/z of 600.
The CTR spectrum above exhibits significant peaks around an m/z of 150 - 200 that have a much lower intensity in the CVD sample. To delve into more details about this specific spectrum, a wide range of functions can be employed:
#' Checking its intensity
intensity(spec2)NumericList of length 1 [[1]] 18.3266733266736 45.1666666666667 … 27.1048951048951 34.9020979020979
#' Checking its rtime
rtime(spec2)[1] 34.872
#' Checking its m/z
mz(spec2)NumericList of length 1 [[1]] 51.1677328505635 53.0461968245186 … 999.139446289161 999.315208803072
#' Filtering for a specific m/z range and viewing in a tabular format
filt_spec <- filterMzRange(spec2,c(50,200))
data.frame(intensity = unlist(intensity(filt_spec)),
mz = unlist(mz(filt_spec))) |>
head() |>
pandoc.table(style = "rmarkdown", caption = "Table 2. Intensity and m/z
values of the 125th spectrum of one CTR sample.")| intensity | mz |
|---|---|
| 18.33 | 51.17 |
| 45.17 | 53.05 |
| 23.08 | 53.7 |
| 41.36 | 53.84 |
| 32.87 | 53.88 |
| 1159 | 54.01 |
Chromatographic Data Visualization: BPC and TIC
The chromatogram() function facilitates the extraction
of intensities along the retention time. However, access to
chromatographic information is currently not as efficient and seamless
as it is for spectral information. Work is underway to develop/improve
the infrastructure for chromatographic data through a new
Chromatograms object aimed to be as flexible and
user-friendly as the Spectra object.
For visualizing LC-MS data, a BPC or TIC serves as a valuable tool to
assess the performance of liquid chromatography across various samples
in an experiment. In our case, we extract the BPC from our data to
create such a plot. The BPC captures the maximum peak signal from each
spectrum in a data file and plots this information against the retention
time for that spectrum on the y-axis. The BPC can be extracted using the
chromatogram function.
By setting the parameter aggregationFun = "max", we
instruct the function to report the maximum signal per spectrum.
Conversely, when setting aggregationFun = "sum", it sums up
all intensities of a spectrum, thereby creating a TIC.
#' Extract and plot BPC for full data
bpc <- chromatogram(data, aggregationFun = "max")
plot(bpc, col = paste0(col_sample, 80), main = "BPC", lwd = 1.5)
grid()
legend("topright", col = col_phenotype,
legend = names(col_phenotype), lty = 1, lwd = 2, horiz = TRUE, bty = "n")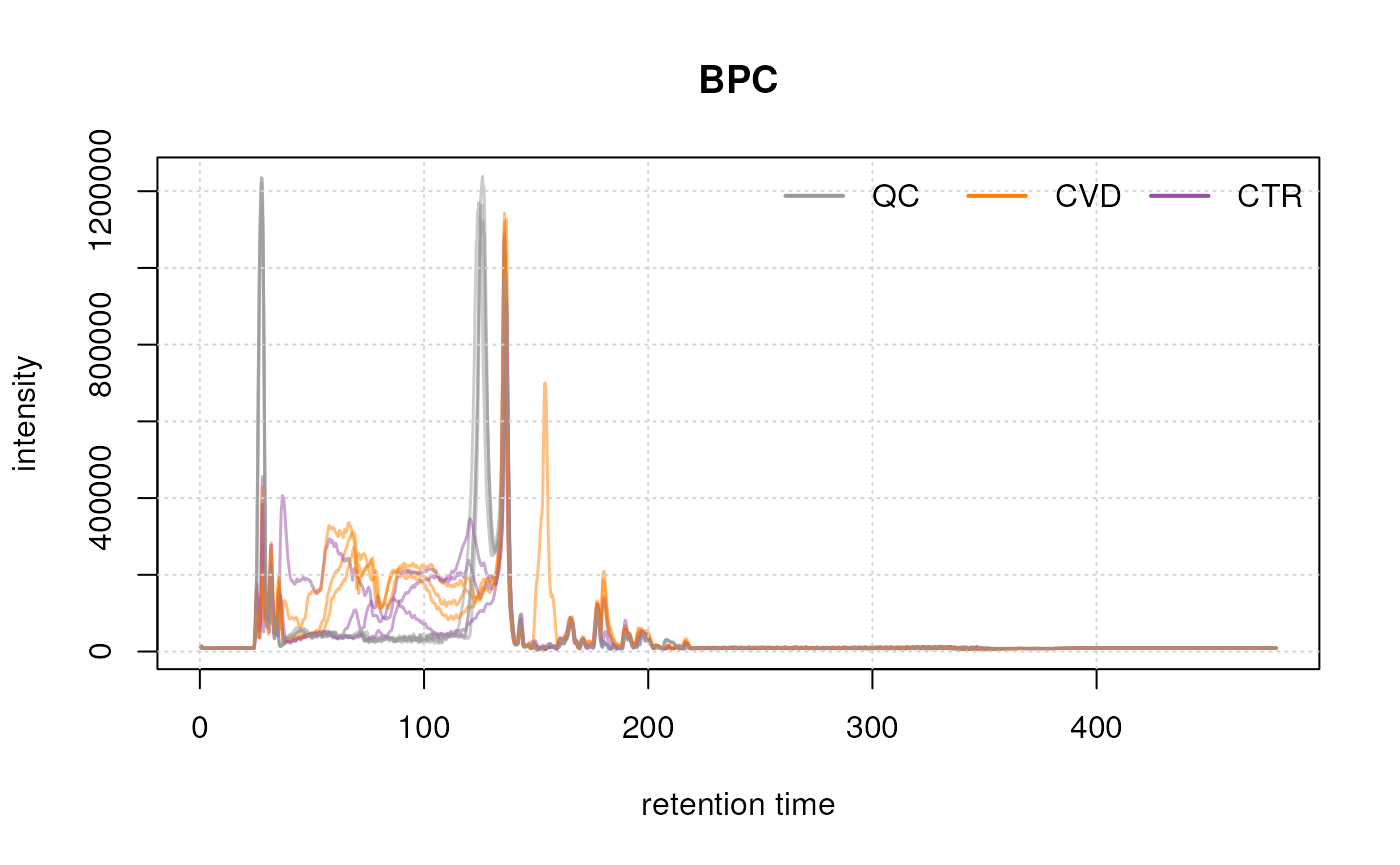
After about 240 seconds no signal seems to be measured. Thus, we filter the data removing that part as well as the first 10 seconds measured in the LC run.
#' Filter the data based on retention time
data <- filterRt(data, c(10, 240))
bpc <- chromatogram(data, aggregationFun = "max")
#' Plot after filtering
plot(bpc, col = paste0(col_sample, 80),
main = "BPC after filtering retention time", lwd = 1.5)
grid()
legend("topright", col = col_phenotype,
legend = names(col_phenotype), lty = 1, lwd = 2, horiz = TRUE, bty = "n")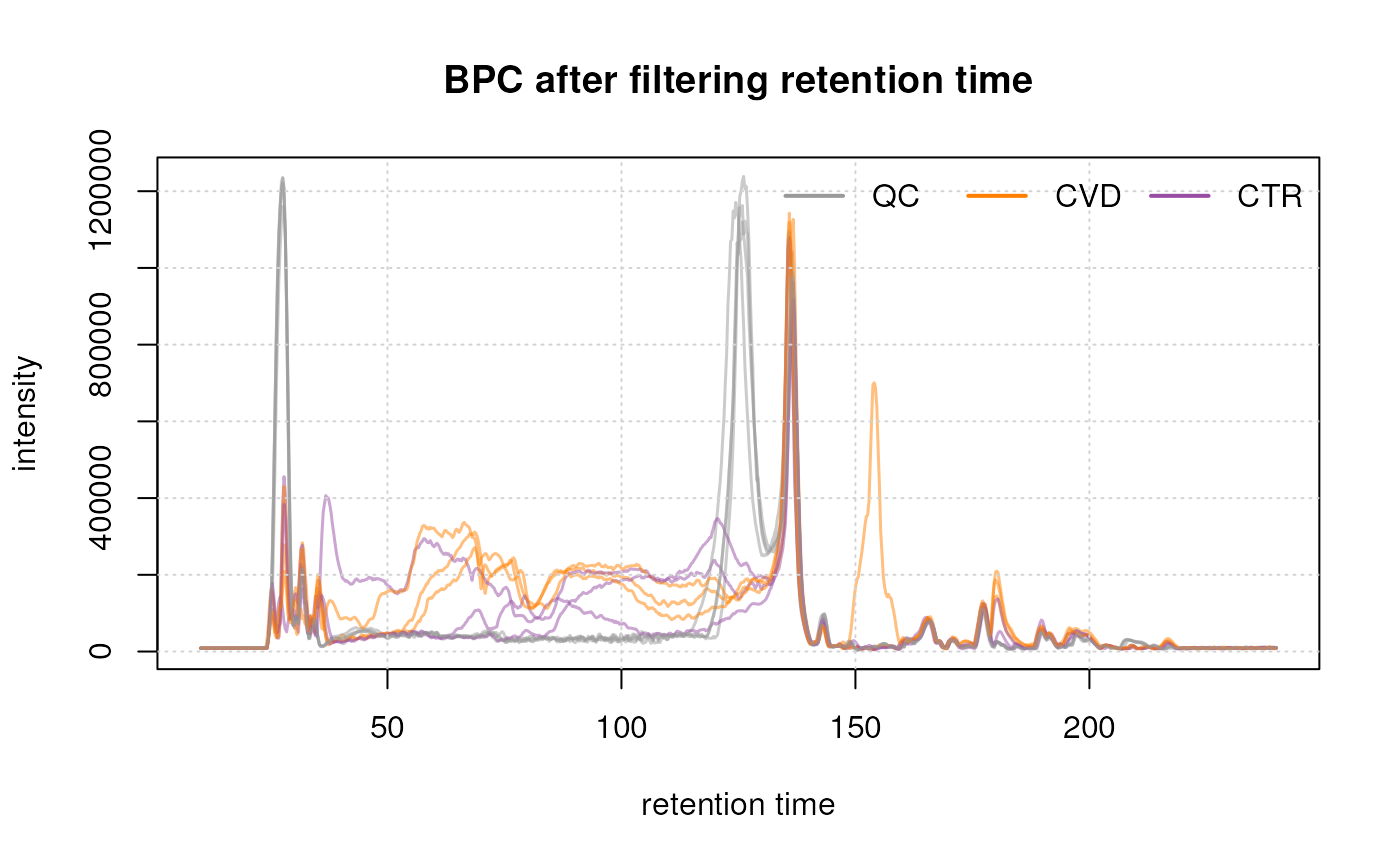
Initially, we examined the entire BPC and subsequently filtered it based on the desired retention times. This not only results in a smaller file size but also facilitates a more straightforward interpretation of the BPC.
The final plot illustrates the BPC for each sample colored by phenotype, providing insights on the signal measured along the retention times of each sample. It reveals the points at which compounds eluted from the LC column. In essence, a BPC condenses the three-dimensional LC-MS data (m/z by retention time by intensity) into two dimensions (retention time by intensity).
We can also here compare similarities of the BPCs in a heatmap. The
retention times will however not be identical between different samples.
Thus we bin() the chromatographic signal per sample along the
retention time axis into bins of two seconds resulting in data with the
same number of bins/data points. We can then calculate pairwise
similarities between these data vectors using the cor()
function and visualize the result using pheatmap().
#' Total ion chromatogram
tic <- chromatogram(data, aggregationFun = "sum") |>
bin(binSize = 2)
#' Calculate similarity (Pearson correlation) between BPCs
ticmap <- do.call(cbind, lapply(tic, intensity)) |>
cor()
rownames(ticmap) <- colnames(ticmap) <- sampleData(data)$sample_name
ann <- data.frame(phenotype = sampleData(data)[, "phenotype"])
rownames(ann) <- rownames(ticmap)
#' Plot heatmap
pheatmap(ticmap, annotation_col = ann,
annotation_colors = list(phenotype = col_phenotype))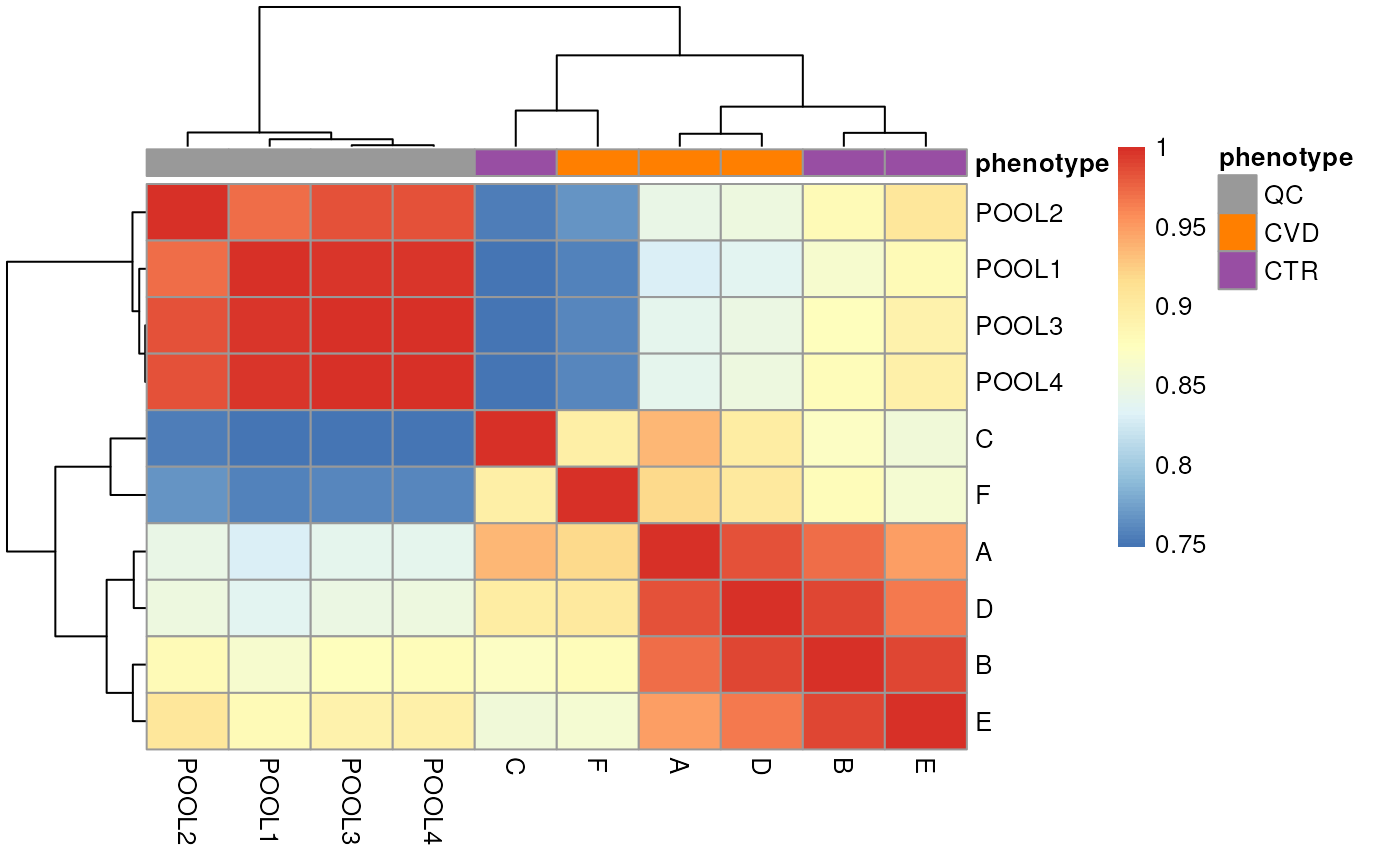
The heatmap above reinforces what our exploration of spectra data showed, which is a strong separation between the QC and study samples. This is important to bear in mind for later analyses.
Additionally, study samples group into two clusters, cluster I containing samples C and F and cluster II with all other samples. Below we plot the TIC of all samples, using a different color for each cluster.
cluster_I_idx <- sampleData(data)$sample_name %in% c("F", "C")
cluster_II_idx <- sampleData(data)$sample_name %in% c("A", "B", "D", "E")
temp_col <- c("grey", "red")
names(temp_col) <- c("Cluster II", "Cluster I")
col <- rep(temp_col[1], length(data))
col[cluster_I_idx] <- temp_col[2]
col[sampleData(data)$phenotype == "QC"] <- NA
data |>
chromatogram(aggregationFun = "sum") |>
plot( col = col,
main = "TIC after filtering retention time", lwd = 1.5)
grid()
legend("topright", col = temp_col,
legend = names(temp_col), lty = 1, lwd = 2,
horiz = TRUE, bty = "n")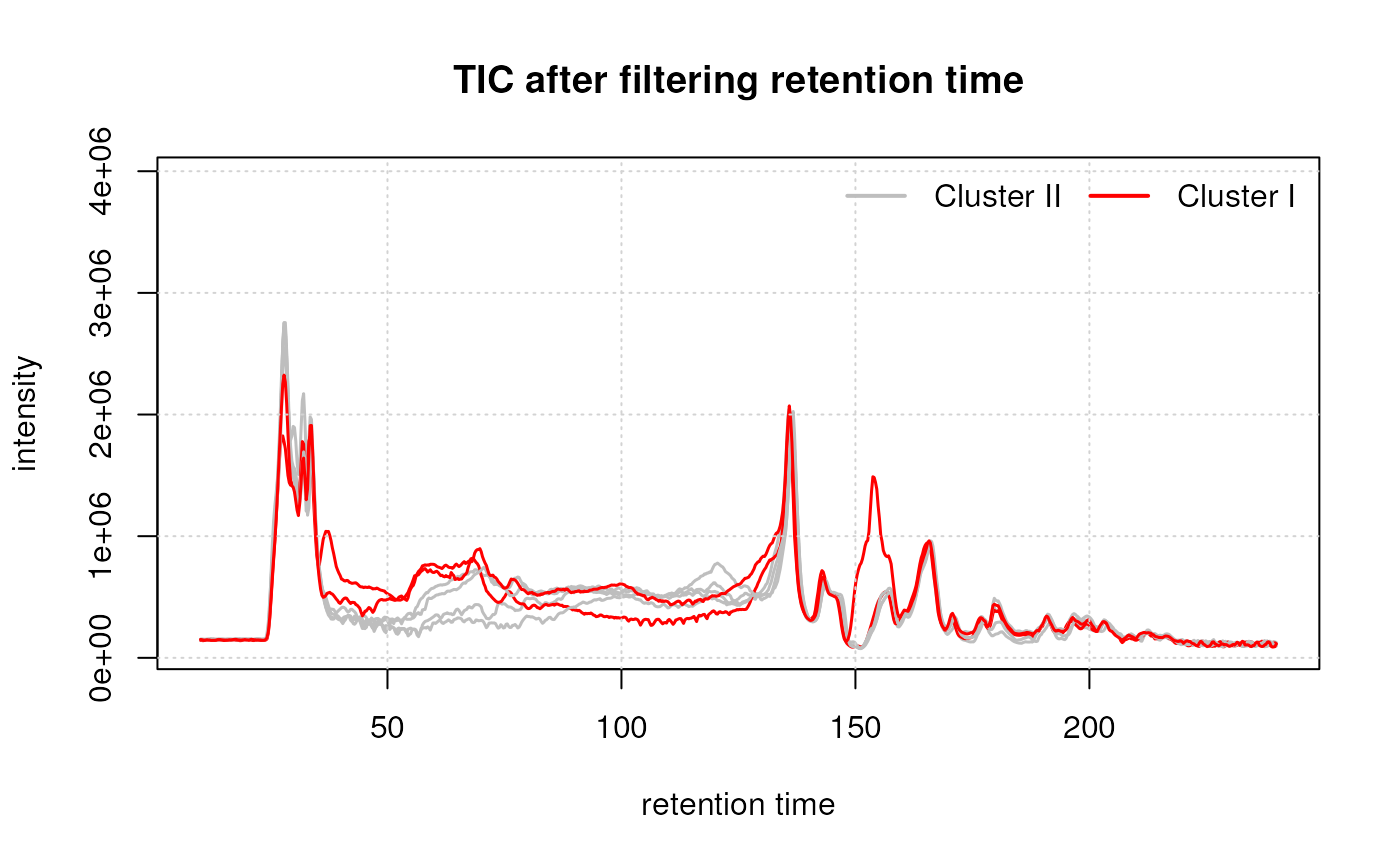
While the TIC of all samples look similar, the samples from cluster I show a different signal in the retention time range from about 40 to 160 seconds. Whether, and how strong this difference will impact the following analysis remains to be determined.
Known compounds
Throughout the entire process, it is crucial to have reference points within the dataset, such as well-known ions. Most experiments nowadays include internal standards (IS), and it was the case here. We strongly recommend using them for visualization throughout the entire analysis. For this experiment, a set of 15 IS was spiked to all samples. After reviewing signal of all of them, we selected two to guide this analysis process. However, we also advise to plot and evaluate all the ions after each steps.
To illustrate this, we generate Extracted Ion Chromatograms (EIC) for these selected test ions. By restricting the MS data to intensities within a restricted, small m/z range and a selected retention time window, EICs are expected to contain only signal from a single type of ion. The expected m/z and retention times for our set of IS was determined in a different experiment. Additionally, in cases where internal standards are not available, commonly present ions in the sample matrix can serve as suitable alternatives. Ideally, these compounds should be distributed across the entire retention time range of the experiment.
#' Load our list of standard
intern_standard <- read.delim("intern_standard_list.txt")
# Extract EICs for the list
eic_is <- chromatogram(
data,
rt = as.matrix(intern_standard[, c("rtmin", "rtmax")]),
mz = as.matrix(intern_standard[, c("mzmin", "mzmax")]))
#' Add internal standard metadata
fData(eic_is)$mz <- intern_standard$mz
fData(eic_is)$rt <- intern_standard$RT
fData(eic_is)$name <- intern_standard$name
fData(eic_is)$abbreviation <- intern_standard$abbreviation
rownames(fData(eic_is)) <- intern_standard$abbreviation
#' Summary of IS information
cpt <- paste("Table 3.Internal standard list with respective m/z and expected",
"retention time [s].")
fData(eic_is)[, c("name", "mz", "rt")] |>
as.data.frame() |>
pandoc.table(style = "rmarkdown", caption = cpt)| name | mz | |
|---|---|---|
| carnitine_d3 | Carnitine (D3) | 165.1 |
| creatinine_methyl_d3 | Creatinine (N-Methhyl_D3) | 117.1 |
| glucose_d2 | Glucose (6,6-D2) | 205.1 |
| alanine_13C_15N | L-Alanine (13C3, 99%; 15N, 99%) | 94.06 |
| arginine_13C_15N | L-Arginine HCl (13C6, 99%; 15N4, 99%) | 185.1 |
| aspartic_13C_15N | L-Aspartic acid (13C4, 99%; 15N, 99%) | 139.1 |
| cystine_13C_15N | L-Cystine (13C6, 99%; 15N2, 99%) | 249 |
| glutamic_13C_15N | L-Glutamic acid (13C5, 99%; 15N, 99%) | 154.1 |
| glycine_13C_15N | Glycine (13C2, 99%; 15N, 99%) | 79.04 |
| histidine_13C_15N | L-Histidine HCl H2O (13C6; 15N3, 99%) | 165.1 |
| isoleucine_13C_15N | L-Isoleucine (13C6, 99%; 15N, 99%) | 139.1 |
| leucine_13C_15N | L-Leucine (13C6, 99%; 15N, 99%) | 139.1 |
| lysine_13C_15N | L-Lysine 2HCl (13C6, 99%; 15N2, 99%) | 155.1 |
| methionine_13C_15N | L-Methionine (13C5, 99%; 15N, 99%) | 156.1 |
| phenylalanine_13C_15N | L-Phenylalanine (13C9, 99%; 15N, 99%) | 176.1 |
| proline_13C_15N | L-Proline (13C5, 99%; 15N, 99%) | 122.1 |
| serine_13C_15N | L-Serine (13C3, 99%; 15N, 99%) | 110.1 |
| threonine_13C_15N | L-Threonine (13C4, 99%; 15N, 99%) | 125.1 |
| tyrosine_13C_15N | L-Tyrosine (13C9, 99%; 15N, 99%) | 192.1 |
| valine_13C_15N | L-Valine (13C5, 99%; 15N, 99%) | 124.1 |
| rt | |
|---|---|
| carnitine_d3 | 61 |
| creatinine_methyl_d3 | 126 |
| glucose_d2 | 166 |
| alanine_13C_15N | 167 |
| arginine_13C_15N | 183 |
| aspartic_13C_15N | 179 |
| cystine_13C_15N | 209 |
| glutamic_13C_15N | 171 |
| glycine_13C_15N | 168 |
| histidine_13C_15N | 185 |
| isoleucine_13C_15N | 154 |
| leucine_13C_15N | 151 |
| lysine_13C_15N | 184 |
| methionine_13C_15N | 161 |
| phenylalanine_13C_15N | 153 |
| proline_13C_15N | 167 |
| serine_13C_15N | 178 |
| threonine_13C_15N | 171 |
| tyrosine_13C_15N | 166 |
| valine_13C_15N | 164 |
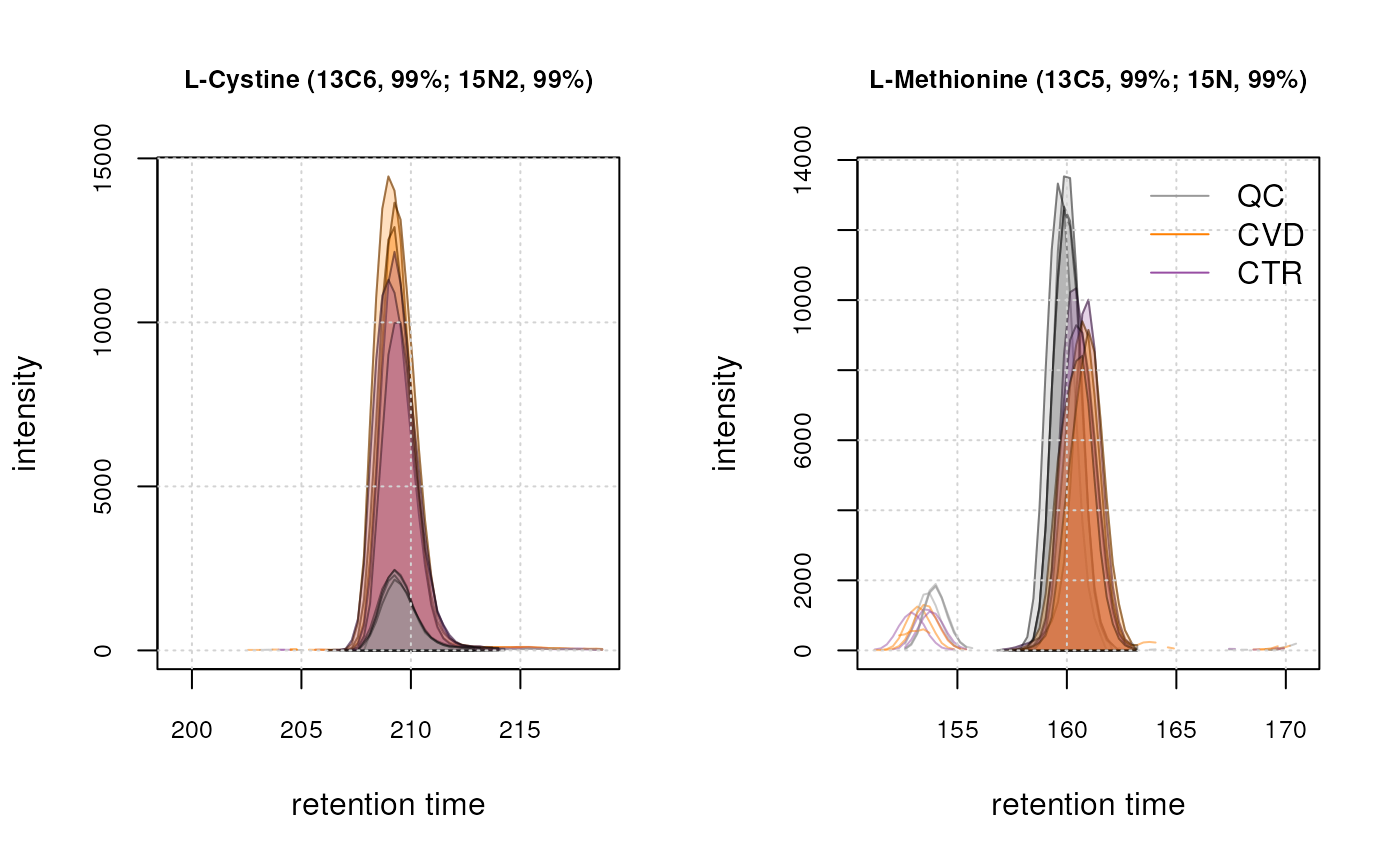
We below plot the EICs for isotope labeled cystine and methionine.
#' Extract the two IS from the chromatogram object.
eic_cystine <- eic_is["cystine_13C_15N"]
eic_met <- eic_is["methionine_13C_15N"]
#' plot both EIC
par(mfrow = c(1, 2), mar = c(4, 2, 2, 0.5))
plot(eic_cystine, main = fData(eic_cystine)$name, cex.axis = 0.8,
cex.main = 0.8,
col = paste0(col_sample, 80))
grid()
abline(v = fData(eic_cystine)$rt, col = "red", lty = 3)
plot(eic_met, main = fData(eic_met)$name, cex.axis = 0.8, cex.main = 0.8,
col = paste0(col_sample, 80))
grid()
abline(v = fData(eic_met)$rt, col = "red", lty = 3)
legend("topright", col = col_phenotype, legend = names(col_phenotype), lty = 1,
bty = "n")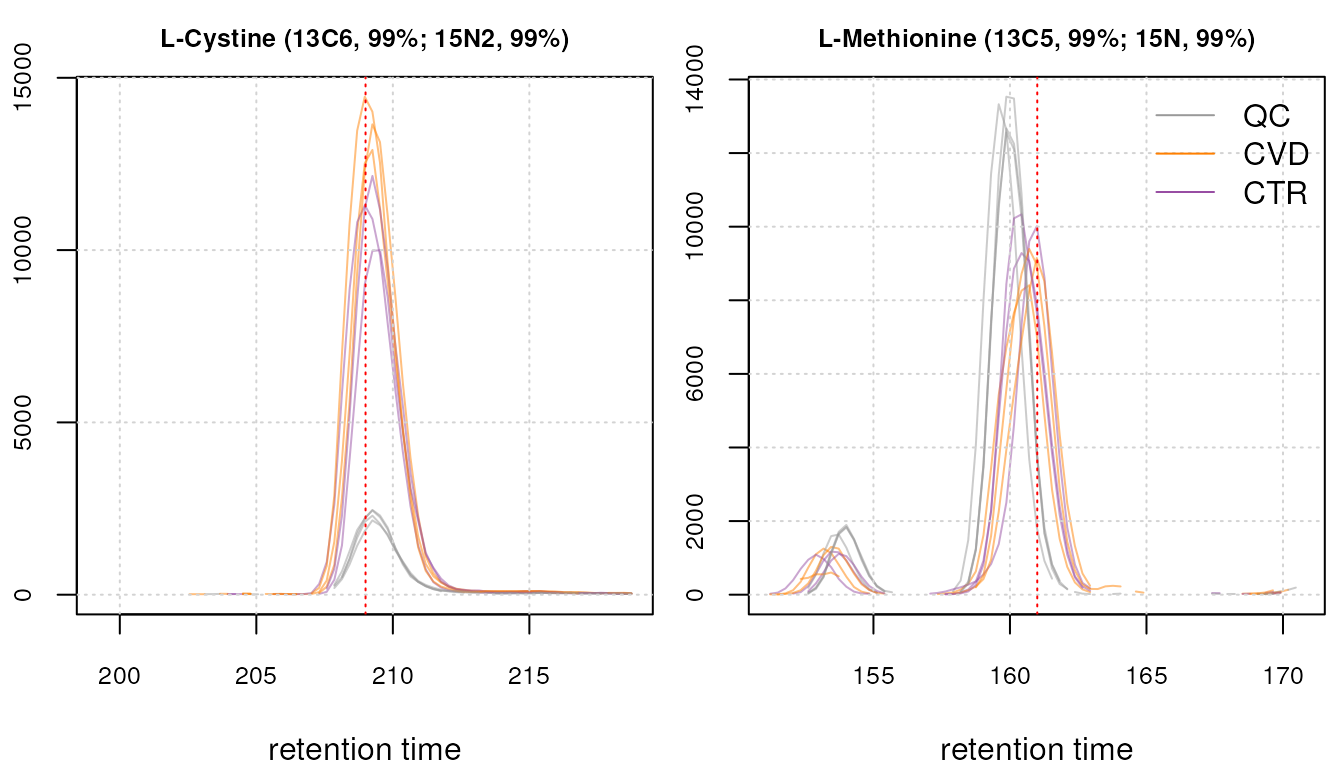
We can observe a clear concentration difference between QCs and study samples for the isotope labeled cystine ion. Meanwhile, the labeled methionine internal standard exhibits a discernible signal amidst some noise and a noticeable retention time shift between samples.
While the artificially isotope labeled compounds were spiked to the
individual samples, there should also be the signal from the endogenous
compounds in serum (or plasma) samples. Thus, we calculate next the mass
and m/z of an [M+H]+ ion of the endogenous cystine
from its chemical formula and extract also the EIC from this ion. For
calculation of the exact mass and the m/z of the selected ion
adduct we use the calculateMass() and
mass2mz() functions from the
r Biocpkg("MetaboCoreUtils") package.
#' extract endogenous cystine mass and EIC and plot.
cysmass <- calculateMass("C6H12N2O4S2")
cys_endo <- mass2mz(cysmass, adduct = "[M+H]+")[, 1]
#' Plot versus spiked
par(mfrow = c(1, 2))
chromatogram(data, mz = cys_endo + c(-0.005, 0.005),
rt = unlist(fData(eic_cystine)[, c("rtmin", "rtmax")]),
aggregationFun = "max") |>
plot(col = paste0(col_sample, 80)) |>
grid()
plot(eic_cystine, col = paste0(col_sample, 80))
grid()
legend("topright", col = col_phenotype, legend = names(col_phenotype), lty = 1,
bty = "n")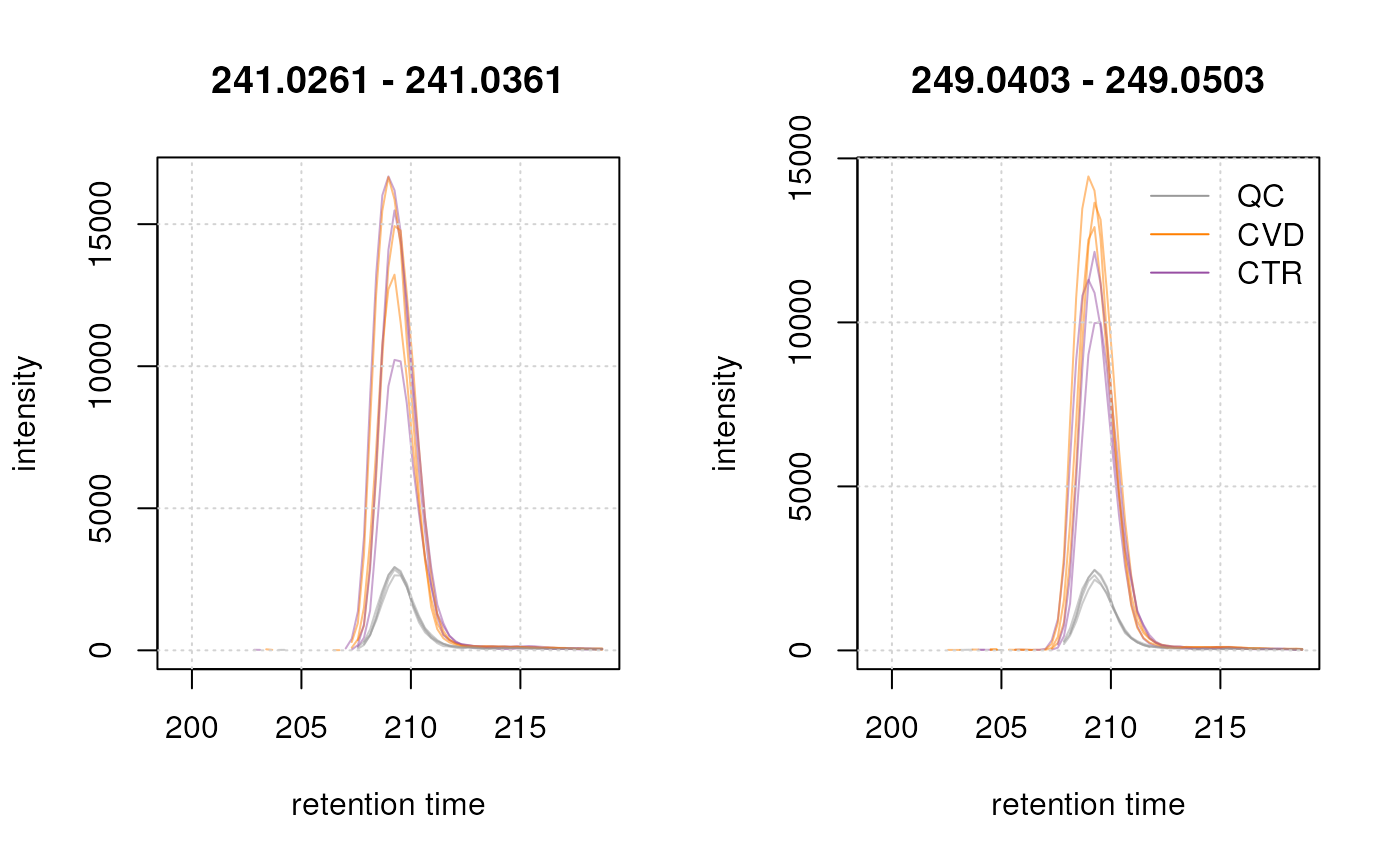
The two cystine EICs above look highly similar (the endogenous shown left, the isotope labeled right in the plot above), if not for the shift in m/z, which arises from the artificial labeling. This shift allows us to discriminate between the endogenous and non-endogenous compound.
Data preprocessing
Preprocessing stands as the inaugural step in the analysis of untargeted LC-MS. It is characterized by 3 main stages: chromatographic peak detection, retention time shift correction (alignment) and correspondence which results in features defined. The primary objective of preprocessing is the quantification of signals from ions measured in a sample, addressing any potential retention time drifts between samples, and ensuring alignment of quantified signals across samples within an experiment. The final result of LC-MS data preprocessing is a numeric matrix with abundances of quantified entities in the samples of the experiment.
[anna: silly question: isn’t the goal of preprocessing to align and group all signals pertaining to a certain ion in a feature? And then obtain the matrix with the abundances][phili: I actually really like anna’s simple definition. what do you think ?]
Chromatographic peak detection
The initial preprocessing step involves detecting intensity peaks
along the retention time axis, the so called chromatographic
peaks. To achieve this, we employ the findChromPeaks()
function within xcms. This function supports various algorithms
for peak detection, which can be selected and configured with their
respective parameter objects.
The preferred algorithm in this case, CentWave, utilizes continuous wavelet transformation (CWT)-based peak detection (Tautenhahn, Böttcher, and Neumann 2008). This method is known for its effectiveness in handling non-Gaussian shaped chromatographic peaks or peaks with varying retention time widths, which are commonly encountered in HILIC separations.
Below we apply the CentWave algorithm with its default settings on the extracted ion chromatogram for cystine and methionine ions and evaluate its results.
#' Use default Centwave parameter
param <- CentWaveParam()
#' Look at the default parameters
param## Object of class: CentWaveParam
## Parameters:
## - ppm: [1] 25
## - peakwidth: [1] 20 50
## - snthresh: [1] 10
## - prefilter: [1] 3 100
## - mzCenterFun: [1] "wMean"
## - integrate: [1] 1
## - mzdiff: [1] -0.001
## - fitgauss: [1] FALSE
## - noise: [1] 0
## - verboseColumns: [1] FALSE
## - roiList: list()
## - firstBaselineCheck: [1] TRUE
## - roiScales: numeric(0)
## - extendLengthMSW: [1] FALSE
## - verboseBetaColumns: [1] FALSE
#' Evaluate for Cystine
cystine_test <- findChromPeaks(eic_cystine, param = param)
chromPeaks(cystine_test)## rt rtmin rtmax into intb maxo sn row column
#' Evaluate for Methionine
met_test <- findChromPeaks(eic_met, param = param)
chromPeaks(met_test)## rt rtmin rtmax into intb maxo sn row columnWhile CentWave is a highly performant algorithm, it requires to be costumized to each dataset. This implies that the parameters should be fine-tuned based on the user’s data. The example above serves as a clear motivation for users to familiarize themselves with the various parameters and the need to adapt them to a data set. We will discuss the main parameters that can be easily adjusted to suit the user’s dataset:
peakwidth: Specifies the minimal and maximal expected width of the peaks in the retention time dimension. Highly dependent on the chromatographic settings used.ppm: The maximal allowed difference of mass peaks’ m/z values (in parts-per-million) in consecutive scans to consider them representing signal from the same ion.integrate: This parameter defines the integration method. Here, we primarily useintegrate = 2because it integrates also signal of a chromatographic peak’s tail and is considered more accurate by the developers.
To determine peakwidth, we recommend that users refer to
previous EICs and estimate the range of peak widths they observe in
their dataset. Ideally, examining multiple EICs should be the goal. For
this dataset, the peak widths appear to be around 2 to 10 seconds. We do
not advise choosing a range that is too wide or too narrow with the
peakwidth parameter as it can lead to false positives or
negatives.
To determine the ppm, a deeper analysis of the dataset
is needed. It is clarified above that ppm depends on the
instrument, but users should not necessarily input the vendor-advertised
ppm. Here’s how to determine it as accurately as possible:
The following steps involve generating a highly restricted MS area with a single mass peak per spectrum, representing the cystine ion. The m/z of these peaks is then extracted, their absolute difference calculated and finally expressed in ppm.
#' Restrict the data to signal from cystine in the first sample
cst <- data[1L] |>
spectra() |>
filterRt(rt = c(208, 218)) |>
filterMzRange(mz = fData(eic_cystine)["cystine_13C_15N", c("mzmin", "mzmax")])
#' Show the number of peaks per m/z filtered spectra
lengths(cst)## [1] 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 0 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1
#' Calculate the difference in m/z values between scans
mz_diff <- cst |>
mz() |>
unlist() |>
diff() |>
abs()
#' Express differences in ppm
range(mz_diff * 1e6 / mean(unlist(mz(cst))))## [1] 0.08829605 14.82188728We therefore, choose a value close to the maximum within this range
for parameter ppm, i.e., 15 ppm.
We can now perform the chromatographic peak detection with the
adapted settings on our EICs. It is important to note that, to properly
estimate background noise, sufficient data points outside the
chromatographic peak need to be present. This is generally no problem if
peak detection is performed on the full LC-MS data set, but for peak
detection on EICs the retention time range of the EIC needs to be
sufficiently wide. If the function fails to find a peak in an EIC, the
initial troubleshooting step should be to increase this range.
Additionally, the signal-to-noise threshold snthresh should
be reduced for peak detection in EICs, because within the small
retention time range, not enough signal is present to properly estimate
the background noise. Finally, in case of too few MS1 data points per
peaks, setting CentWave’s advanced parameter
extendLengthMSW to TRUE can help with peak
detection.
#' Parameters adapted for chromatographic peak detection on EICs.
param <- CentWaveParam(peakwidth = c(1, 8), ppm = 15, integrate = 2,
snthresh = 2)
#' Evaluate on the cystine ion
cystine_test <- findChromPeaks(eic_cystine, param = param)
chromPeaks(cystine_test)## rt rtmin rtmax into intb maxo sn row column
## [1,] 209.251 207.577 212.878 4085.675 2911.376 2157.459 4 1 1
## [2,] 209.251 206.182 213.995 24625.728 19074.407 12907.487 4 1 2
## [3,] 209.252 207.020 214.274 19467.836 14594.041 9996.466 4 1 3
## [4,] 209.251 207.577 212.041 4648.229 3202.617 2458.485 3 1 4
## [5,] 208.974 206.184 213.159 23801.825 18126.978 11300.289 3 1 5
## [6,] 209.250 207.018 213.714 25990.327 21036.768 13650.329 5 1 6
## [7,] 209.252 207.857 212.879 4528.767 3259.039 2445.841 4 1 7
## [8,] 209.252 207.299 213.995 23119.449 17274.140 12153.410 4 1 8
## [9,] 208.972 206.740 212.878 28943.188 23436.119 14451.023 4 1 9
## [10,] 209.252 207.578 213.437 4470.552 3065.402 2292.881 4 1 10
#' Evaluate on the methionine ion
met_test <- findChromPeaks(eic_met, param = param)
chromPeaks(met_test)## rt rtmin rtmax into intb maxo sn row column
## [1,] 159.867 157.913 162.378 20026.61 14715.42 12555.601 4 1 1
## [2,] 160.425 157.077 163.215 16827.76 11843.39 8407.699 3 1 2
## [3,] 160.425 157.356 163.215 18262.45 12881.67 9283.375 3 1 3
## [4,] 159.588 157.635 161.820 20987.72 15424.25 13327.811 4 1 4
## [5,] 160.985 156.799 163.217 16601.72 11968.46 10012.396 4 1 5
## [6,] 160.982 157.634 163.214 17243.24 12389.94 9150.079 4 1 6
## [7,] 159.867 158.193 162.099 21120.10 16202.05 13531.844 3 1 7
## [8,] 160.426 157.356 162.937 18937.40 13739.73 10336.000 3 1 8
## [9,] 160.704 158.472 163.215 17882.21 12299.43 9395.548 3 1 9
## [10,] 160.146 157.914 162.379 20275.80 14279.50 12669.821 3 1 10With the customized parameters, a chromatographic peak was detected
in each sample. Below, we use the plot() function on the
EICs to visualize these results.
We can see a peak seems ot be detected in each sample for both ions. This indicates that our custom settings seem thus to be suitable for our dataset. We now proceed and apply them to the entire dataset, extracting EICs again for the same ions to evaluate and confirm that chromatographic peak detection worked as expected. Note:
We revert the value for parameter
snthreshto its default, because, as mentioned above, background noise estimation is more reliable when performed on the full data set.Parameter
chunkSizeoffindChromPeaks()defines the number of data files that are loaded into memory and processed simultaneously. This parameter thus allows to fine-tune the memory demand as well as performance of the chromatographic peak detection step.
#' Using the same settings, but with default snthresh
param <- CentWaveParam(peakwidth = c(1, 8), ppm = 15, integrate = 2)
data <- findChromPeaks(data, param = param, chunkSize = 5)
#' Update EIC internal standard object
eics_is_noprocess <- eic_is
eic_is <- chromatogram(data,
rt = as.matrix(intern_standard[, c("rtmin", "rtmax")]),
mz = as.matrix(intern_standard[, c("mzmin", "mzmax")]))
fData(eic_is) <- fData(eics_is_noprocess)Below we plot the EICs of the two selected internal standards to evaluate the chromatographic peak detection results.
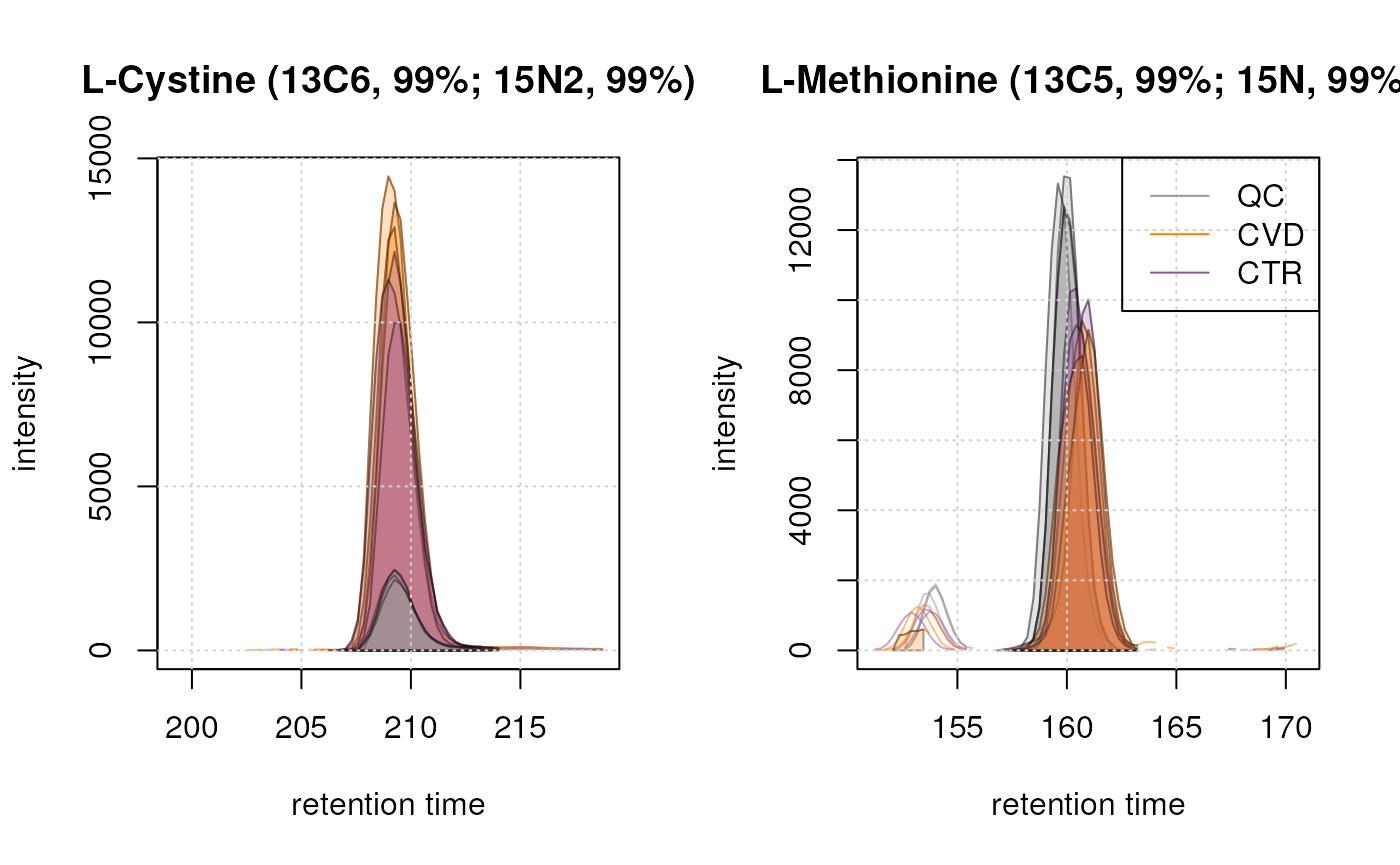
Peaks seem to have been detected properly in all samples for both ions. This indicates that the peak detection process over the entire dataset was successful.
Refine identified chromatographic peaks
The identification of chromatographic peaks using the
CentWave algorithm can sometimes result in artifacts, such as
overlapping or split peaks. To address this issue, the
refineChromPeaks() function is utilized, in conjunction
with MergeNeighboringPeaksParam, which aims at merging such
split peaks.
Below we show some examples of CentWave peak detection artifacts. These examples are pre-selected to illustrate the necessity of the next step:
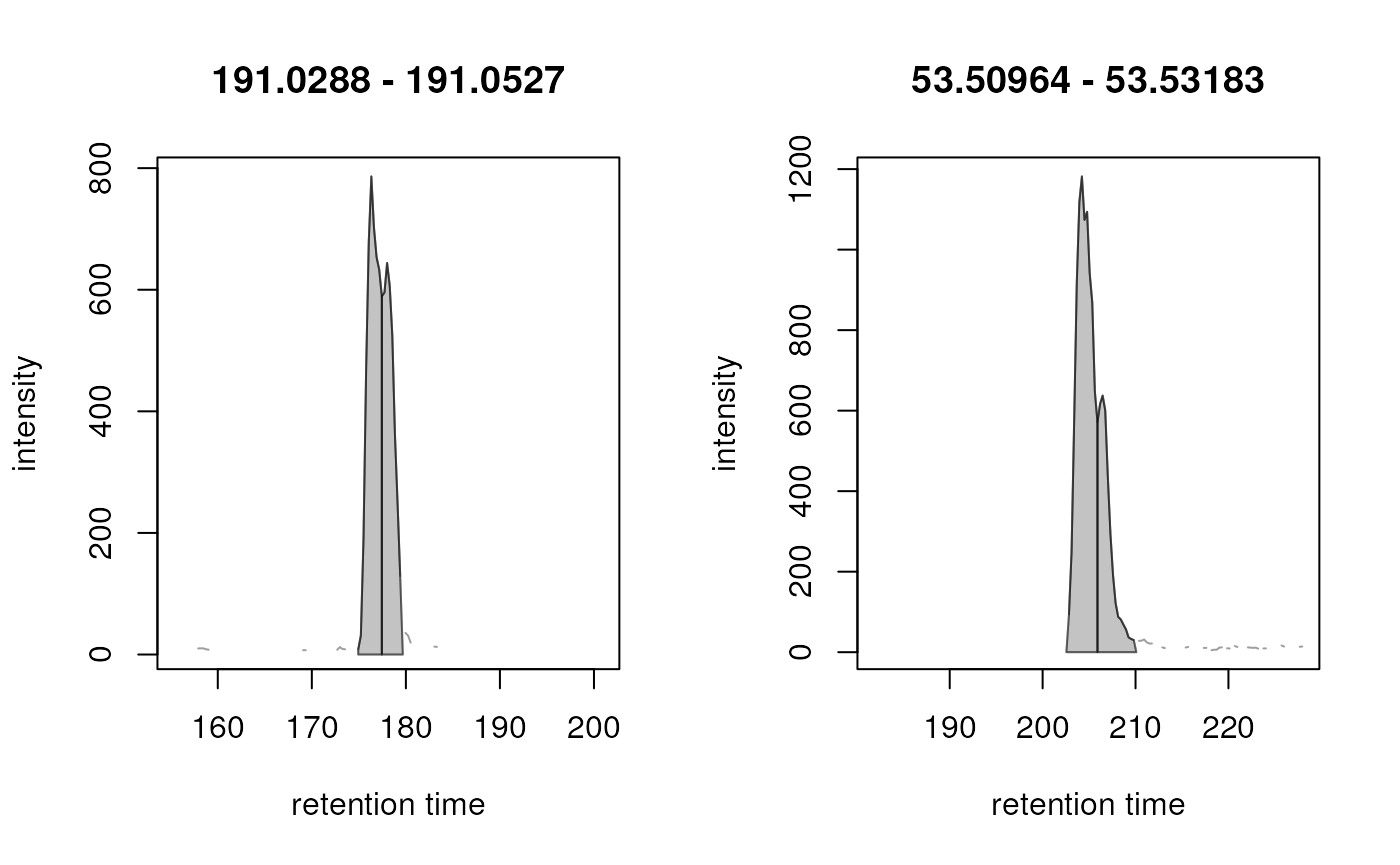
In both cases the signal presumably from a single type of ion was
split into two separate chromatographic peaks (indicated by the vertical
line). The MergeNeigboringPeaksParam allows to combine such
split peaks. The parameters for this algorithm are defined below:
-
expandMzandexpandRt: Define which chromatographic peaks should be evaluated for merging.-
expandMz: Suggested to be kept relatively small (here at 0.0015) to prevent the merging of isotopes. -
expandRt: Usually set to approximately half the size of the average retention time width used for chromatographic peak detection (in this case, 2.5 seconds).
-
-
minProp: Used to determine whether candidates will be merged. Chromatographic peaks with overlapping m/z ranges (expanded on each side byexpandMz) and with a tail-to-head distance in the retention time dimension that is less than2 * expandRt, and for which the signal between them is higher thanminPropof the apex intensity of the chromatographic peak with the lower intensity, are merged. Values for this parameter should not be too small to avoid merging closely co-eluting ions, such as isomers.
We test these settings below on the EICs with the split peaks.
#' set up the parameter
param <- MergeNeighboringPeaksParam(expandRt = 2.5, expandMz = 0.0015,
minProp = 0.75)
#' Perform the peak refinement on the EICs
eics <- refineChromPeaks(eics, param = param)
plot(eics)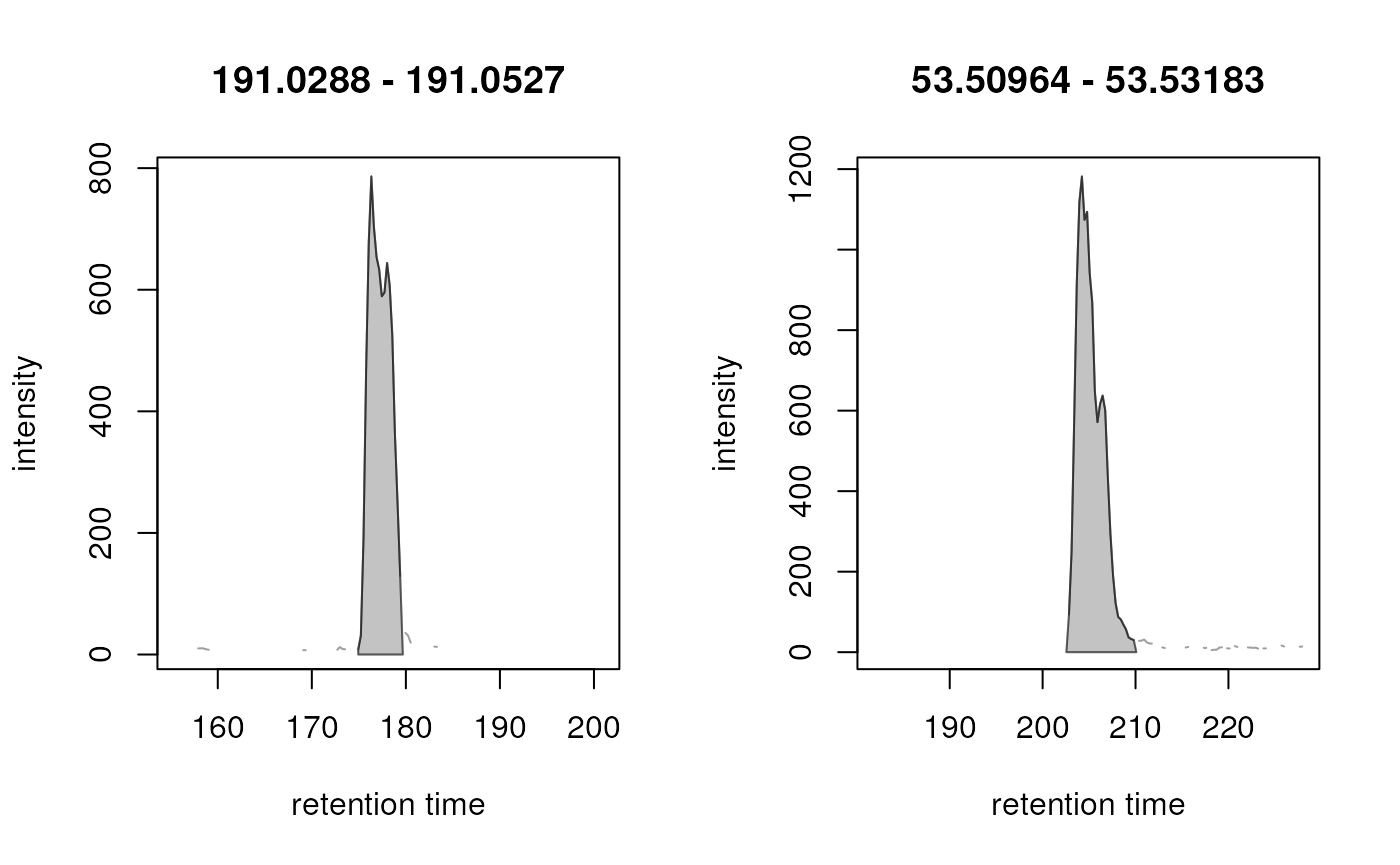
We can observe that the artificially split peaks have been
appropriately merged. Therefore, we next apply these settings to the
entire dataset. After peak merging, column "merged" in the
result object’s chromPeakData() data frame can be used to
evaluate which chromatographic peaks in the result represent signal from
merged, or from the originally identified chromatographic peaks.
#' Apply on whole dataset
data <- refineChromPeaks(data, param = param, chunkSize = 5)
chromPeakData(data)$merged |>
table()##
## FALSE TRUE
## 79908 9274Before proceeding with the next preprocessing step it is generally suggested to evaluate the results of the chromatographic peak detection on EICs of e.g. internal standards or other compounds/ions known to be present in the samples.
eics_is_chrompeaks <- eic_is
eic_is <- chromatogram(data,
rt = as.matrix(intern_standard[, c("rtmin", "rtmax")]),
mz = as.matrix(intern_standard[, c("mzmin", "mzmax")]))
fData(eic_is) <- fData(eics_is_chrompeaks)
eic_cystine <- eic_is["cystine_13C_15N", ]
eic_met <- eic_is["methionine_13C_15N", ]Additionally, evaluating and comparing the number of identified chromatographic peaks in all samples of a data set can help spotting potentially problematic samples. Below we count the number of chromatographic peaks per sample and show these numbers in a table.
#' Count the number of peaks per sample and summarize them in a table.
data.frame(sample_name = sampleData(data)$sample_name,
peak_count = as.integer(table(chromPeaks(data)[, "sample"]))) |>
pandoc.table(
style = "rmarkdown",
caption = "Table 4.Samples and number of identified chromatographic peaks.")| sample_name | peak_count |
|---|---|
| POOL1 | 9287 |
| A | 8986 |
| B | 8738 |
| POOL2 | 9193 |
| C | 8351 |
| D | 8778 |
| POOL3 | 9211 |
| E | 8787 |
| F | 8515 |
| POOL4 | 9336 |
A similar number of chromatographic peaks was identified within the various samples of the data set.
Additional options to evaluate the results of the chromatographic
peak detection can be implemented using the
plotChromPeaks() function or by summarizing the results
using base R commands.
Retention time alignment
Despite using the same chromatographic settings and conditions retention time shifts are unavoidable. Indeed, the performance of the instrument can change over time, for example due to small variations in environmental conditions, such as temperature and pressure. These shifts will be generally small if samples are measured within the same batch/measurement run, but can be considerable if data of an experiment was acquired across a longer time period.
To evaluate the presence of a shift we extract and plot below the BPC from the QC samples.
#' Get QC samples
QC_samples <- sampleData(data)$phenotype == "QC"
#' extract BPC
data[QC_samples] |>
chromatogram(aggregationFun = "max", chromPeaks = "none") |>
plot(col = col_phenotype["QC"], main = "BPC of QC samples") |>
grid()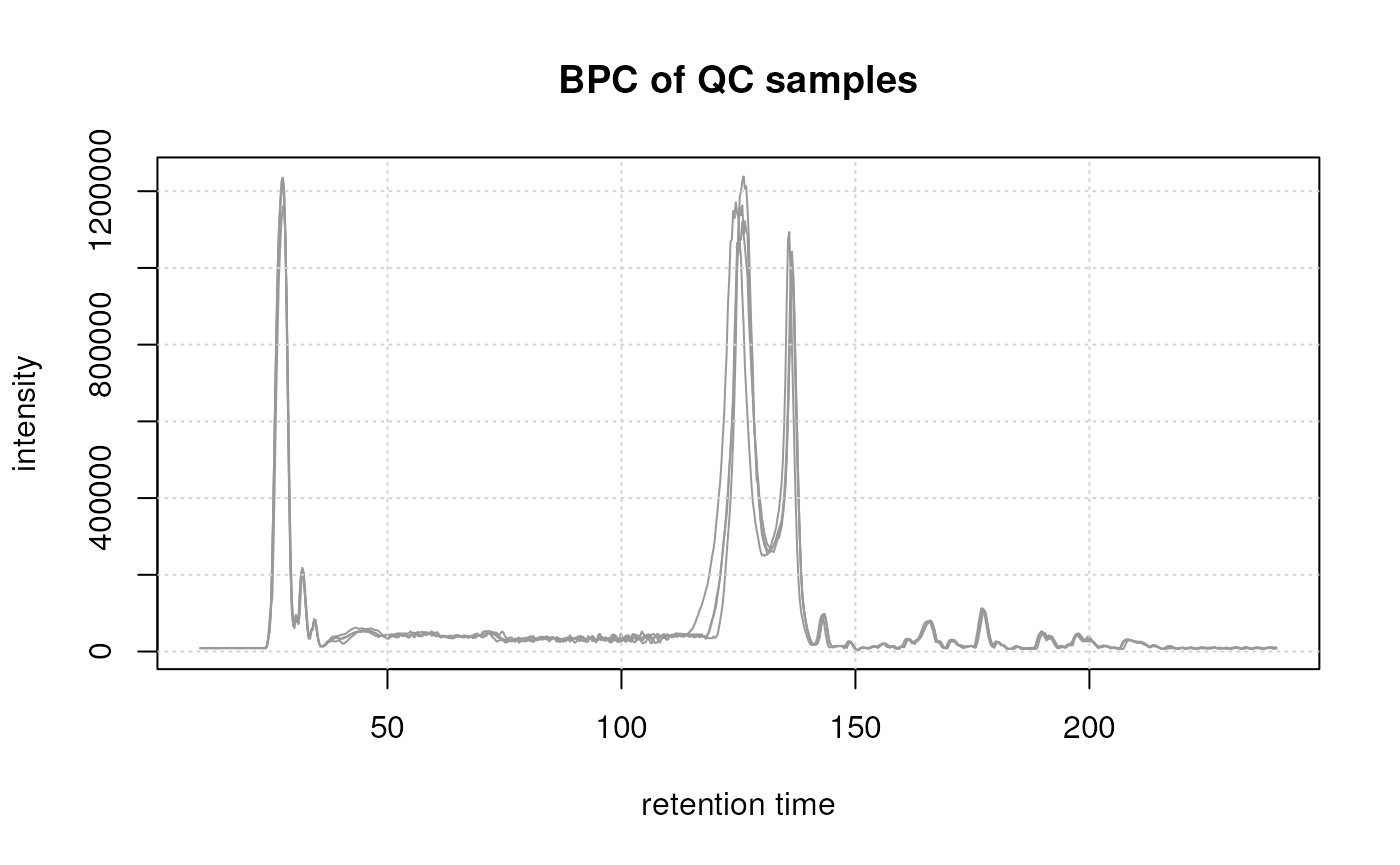
These QC samples representing the same sample (pool) were measured at regular intervals during the measurement run of the experiment and were all measured on the same day. Still, small shifts can be observed, especially in the region between 100 and 150 seconds. To facilitate proper correspondence of signals across samples (and hence definition of the LC-MS features), it is essential to minimize these differences in retention times.
Theoretically, we proceed in two steps: first we select only the QC samples of our dataset and do a first alignment on these, using the so-called anchor peaks. In this way we can assume a linear shift in time, since we are always measuring the same sample in different regular time intervals. Despite having external QCs in our data set, we still use the subset-based alignment assuming retention time shifts to be independent of the different sample matrix (human serum or plasma) and instead are mostly instrument-dependent. Note that it would also be possible to manually specify anchor peaks, respectively their retention times or to align a data set against an external, reference, data set. More information is provided in the vignettes of the xcms package.
After calculating how much to adjust the retention time in these samples, we apply this shift also on the study samples.
In xcms retention time alignment can be performed using the
adjustRtime() function with an alignment algorithm. For
this example we use the PeakGroups method (Smith et al. 2006) that performs the alignment
by minimizing differences in retention times of a set of anchor
peaks in the different samples. This method requires an initial
correspondence analysis to match/group chromatographic peaks across
samples from which the algorithm then selects the anchor peaks for the
alignment.
For the initial correspondence, we use the PeakDensity
approach (Smith et al. 2006) that groups
chromatographic peaks with similar m/z and retention time into
LC-MS features. The parameters for this algorithm, that can be
configured using the PeakDensityParam object, are
sampleGroups, minFraction,
binSize, ppm and bw.
binSize, ppm and bw allow to
specify how similar the chromatographic peaks’ m/z and
retention time values need to be to consider them for grouping into a
feature.
binSizeandppmdefine the required similarity of m/z values. Within each m/z bin (defined bybinSizeandppm) areas along the retention time axis with a high chromatographic peak density (considering all peaks in all samples) are identified, and chromatographic peaks within these regions are considered for grouping into a feature.High density areas are identified using the base R
density()function, for whichbwis a parameter: higher values define wider retention time areas, lower values require chromatographic peaks to have more similar retention times. This parameter can be seen as the black line on the plot below, corresponding to the smoothness of the density curve.
Whether such candidate peaks get grouped into a feature depends also
on parameters sampleGroups and
minFraction:
sampleGroupsshould provide, for each sample, the sample group it belongs to.minFractionis expected to be a value between 0 and 1 defining the proportion of samples within at least one of the sample groups (defined withsampleGroups) in which a chromatographic peaks was detected to group them into a feature.
For the initial correspondence, parameters don’t need to be fully
optimized. Selection of dataset-specific parameter values is described
in more detail in the next section. For our dataset, we use small values
for binSize and ppm and, importantly, also for
parameter bw, since for our data set an ultra high
performance (UHP) LC setup was used [anna: maybe I have been out of this
field for too long, but I don’t see the connection between UHPLC and
choice of small values for these parameters. Is it something empirical?
phili: jo can you help me here ?]. For minFraction we use a
high value (0.9) to ensure only features are defined for chromatographic
peaks present in almost all samples of one sample group (which can then
be used as anchor peaks for the actual alignment). We will base the
alignment later on QC samples only and hence define for
sampleGroups a binary variable grouping samples either into
a study, or QC group.
# Initial correspondence analysis
param <- PeakDensityParam(sampleGroups = sampleData(data)$phenotype == "QC",
minFraction = 0.9,
binSize = 0.01, ppm = 10,
bw = 2)
data <- groupChromPeaks(data, param = param)
plotChromPeakDensity(
eic_cystine, param = param,
col = paste0(col_sample, "80"),
peakCol = col_sample[chromPeaks(eic_cystine)[, "sample"]],
peakBg = paste0(col_sample[chromPeaks(eic_cystine)[, "sample"]], 20),
peakPch = 16)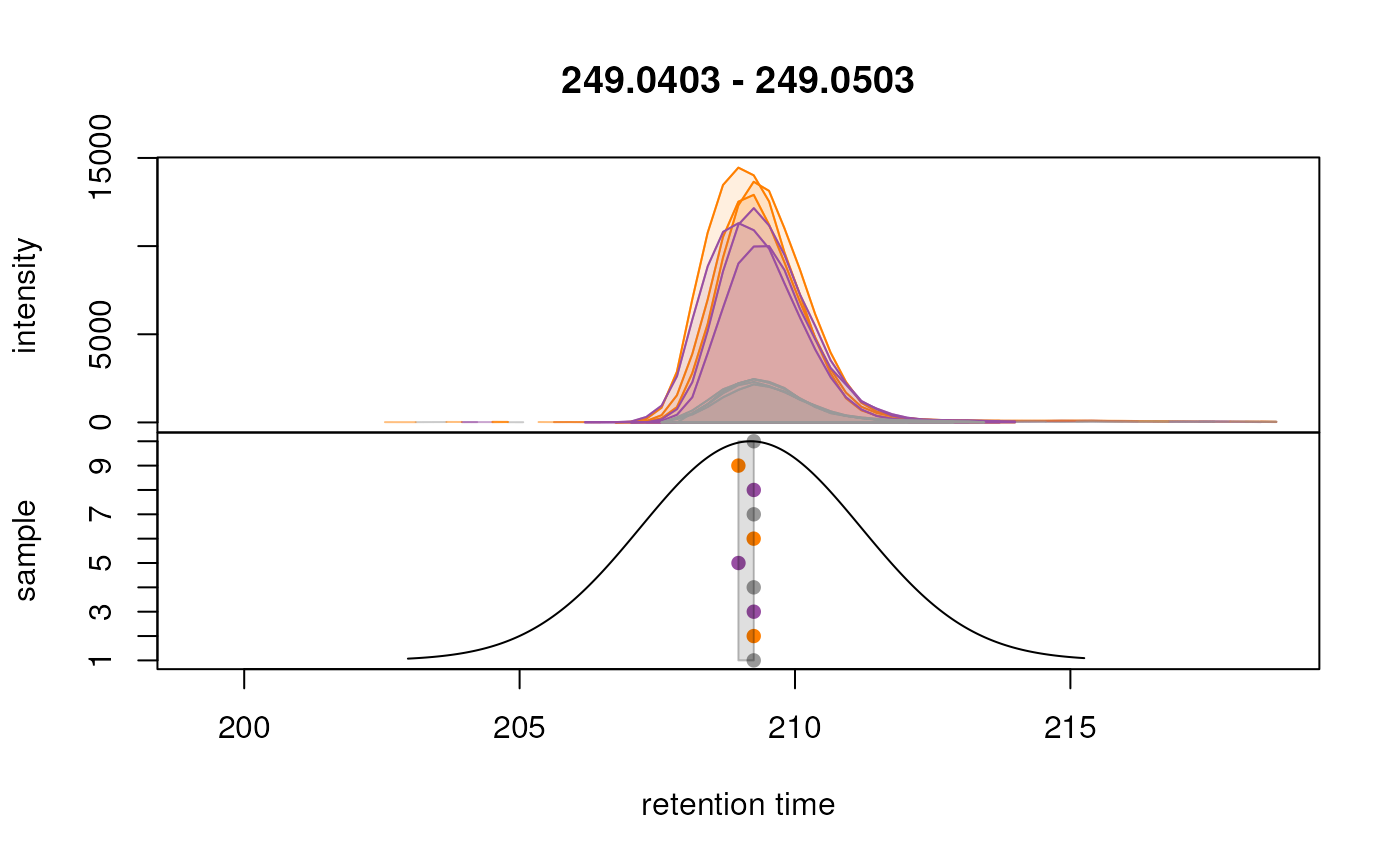
PeakGroups-based alignment can next be performed using the
adjustRtime() function with a PeakGroupsParam
parameter object. The parameters for this algorithm are:
subsetAdjustandsubset: Allows for subset alignment. Here we base the retention time alignment on the QC samples, i.e., retention time shifts will be estimated based on these repeatedly measured samples. The resulting adjustment is then applied to the entire data. For data sets in which QC samples (e.g. sample pools) are measured repeatedly, we strongly suggest to use this method. Note also that for subset-based alignment the samples should be ordered by injection index (i.e., in the order in which they were measured during the measurement run).minFraction: A value between 0 and 1 defining the proportion of samples (of the full data set, or the data subset defined withsubset) in which a chromatographic peak has to be identified to use it as anchor peak. This is in contrast to thePeakDensityParamwhere this parameter was used to define a proportion within a sample group.span: The PeakGroups method allows, depending on the data, to adjust regions along the retention time axis differently. To enable such local alignments the LOESS function is used and this parameter defines the degree of smoothing of this function. Generally, values between 0.4 and 0.6 are used, however, it is suggested to evaluate alignment results and eventually adapt parameters if the result was not satisfactory.
Below we perform the alignment of our data set based on retention times of anchor peaks defined in the subset of QC samples.
#' Define parameters of choice
subset <- which(sampleData(data)$phenotype == "QC")
param <- PeakGroupsParam(minFraction = 0.9, extraPeaks = 50, span = 0.5,
subsetAdjust = "average",
subset = subset)
#' Perform the alignment
data <- adjustRtime(data, param = param)Alignment adjusted the retention times of all spectra in the data set, as well as the retention times of all identified chromatographic peaks.
Once the alignment has been performed, the user should evaluate the
results using the plotAdjustedRtime() function. This
function visualizes the difference between adjusted and raw retention
time for each sample on the y-axis along the adjusted retention time on
the x-axis. Dot points represent the position of the used anchor peak
along the retention time axis. For optimal alignment in all areas along
the retention time axis, these anchor peaks should be scattered all over
the retention time dimension.
#' Visualize alignment results
plotAdjustedRtime(data, col = paste0(col_sample, 80), peakGroupsPch = 1)
grid()
legend("topright", col = col_phenotype,
legend = names(col_phenotype), lty = 1, bty = "n")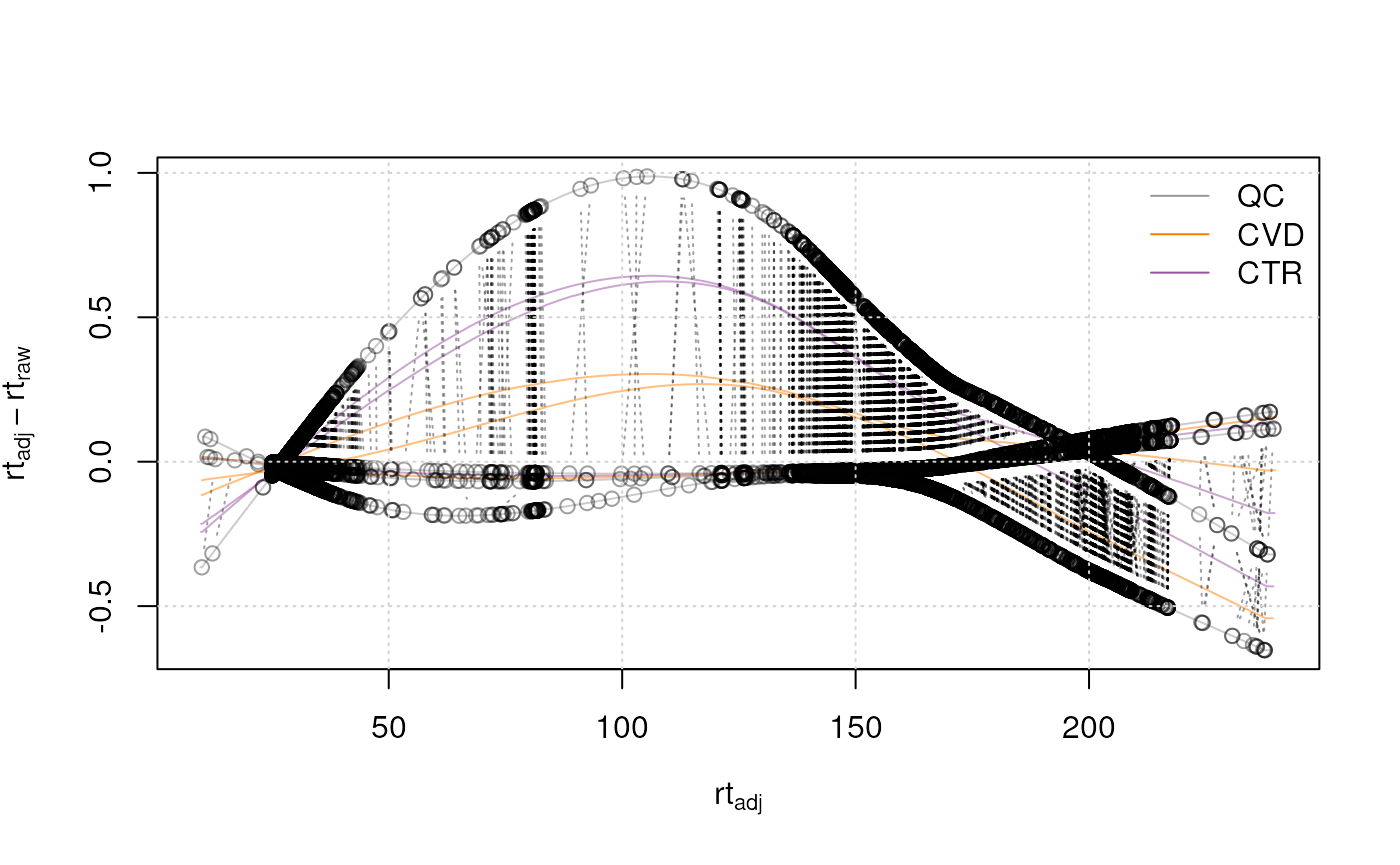
All samples from the present data set were measured within the same measurement run, resulting in small retention time shifts. Therefore, only little adjustments needed to be performed (shifts of at maximum 1 second as can be seen in the plot above). Generally, the magnitude of adjustment seen in such plots should match the expectation from the analyst.
We can also compare the BPC before and after alignment. To get the
original data, i.e. the raw retention times, we can use the
dropAdjustedRtime() function:
#' Get data before alignment
data_raw <- dropAdjustedRtime(data)
#' Apply the adjusted retention time to our dataset
data <- applyAdjustedRtime(data)
#' Plot the BPC before and after alignment
par(mfrow = c(2, 1), mar = c(2, 1, 1, 0.5))
chromatogram(data_raw, aggregationFun = "max", chromPeaks = "none") |>
plot(main = "BPC before alignment", col = paste0(col_sample, 80))
grid()
legend("topright", col = col_phenotype,
legend = names(col_phenotype), lty = 1, bty = "n", horiz = TRUE)
chromatogram(data, aggregationFun = "max", chromPeaks = "none") |>
plot(main = "BPC after alignment",
col = paste0(col_sample, 80))
grid()
legend("topright", col = col_phenotype,
legend = names(col_phenotype), lty = 1, bty = "n", horiz = TRUE)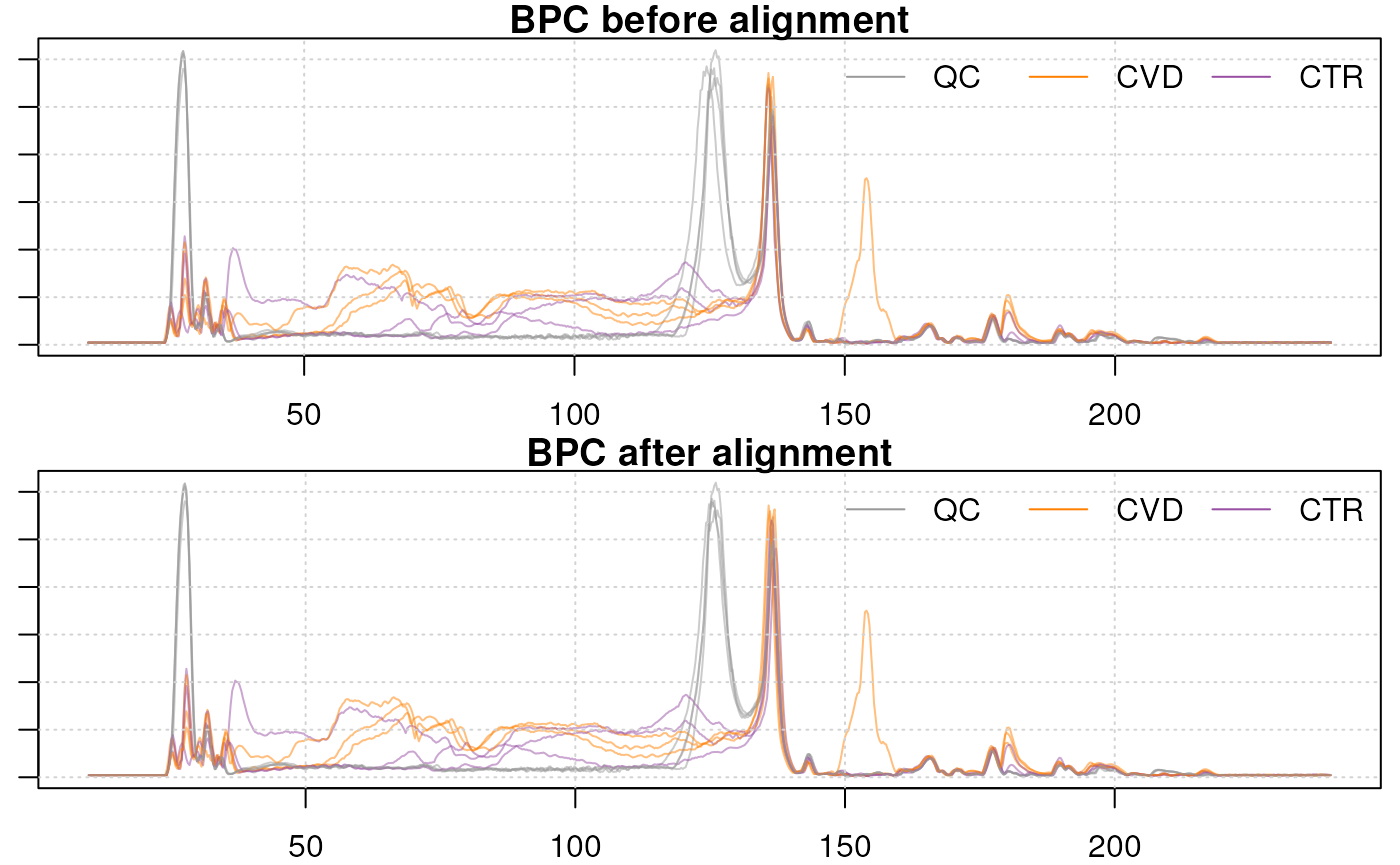
The largest shift can be observed in the retention time range from 120 to 130s. Apart from that retention time range, only little changes can be observed.
We next evaluate the impact of the alignment on the EICs of the selected internal standards. We thus below first extract the ion chromatograms after alignment.
#' Store the EICs before alignment
eics_is_refined <- eic_is
#' Update the EICs
eic_is <- chromatogram(data,
rt = as.matrix(intern_standard[, c("rtmin", "rtmax")]),
mz = as.matrix(intern_standard[, c("mzmin", "mzmax")]))
fData(eic_is) <- fData(eics_is_refined)
#' Extract the EICs for the test ions
eic_cystine <- eic_is["cystine_13C_15N"]
eic_met <- eic_is["methionine_13C_15N"]We can now evaluate the alignment effect in our test ions. We will plot the EICs before and after alignment for both the isotope labeled cystine and methionine.
par(mfrow = c(2, 2), mar = c(4, 4.5, 2, 1))
old_eic_cystine <- eics_is_refined["cystine_13C_15N"]
plot(old_eic_cystine, main = "Cystine before alignment", peakType = "none",
col = paste0(col_sample, 80))
grid()
abline(v = intern_standard["cystine_13C_15N", "RT"], col = "red", lty = 3)
old_eic_met <- eics_is_refined["methionine_13C_15N"]
plot(old_eic_met, main = "Methionine before alignment",
peakType = "none", col = paste0(col_sample, 80))
grid()
abline(v = intern_standard["methionine_13C_15N", "RT"], col = "red", lty = 3)
plot(eic_cystine, main = "Cystine after alignment", peakType = "none",
col = paste0(col_sample, 80))
grid()
abline(v = intern_standard["cystine_13C_15N", "RT"], col = "red", lty = 3)
legend("topright", col = col_phenotype,
legend = names(col_phenotype), lty = 1, bty = "n")
plot(eic_met, main = "Methionine after alignment",
peakType = "none", col = paste0(col_sample, 80))
grid()
abline(v = intern_standard["methionine_13C_15N", "RT"], col = "red", lty = 3)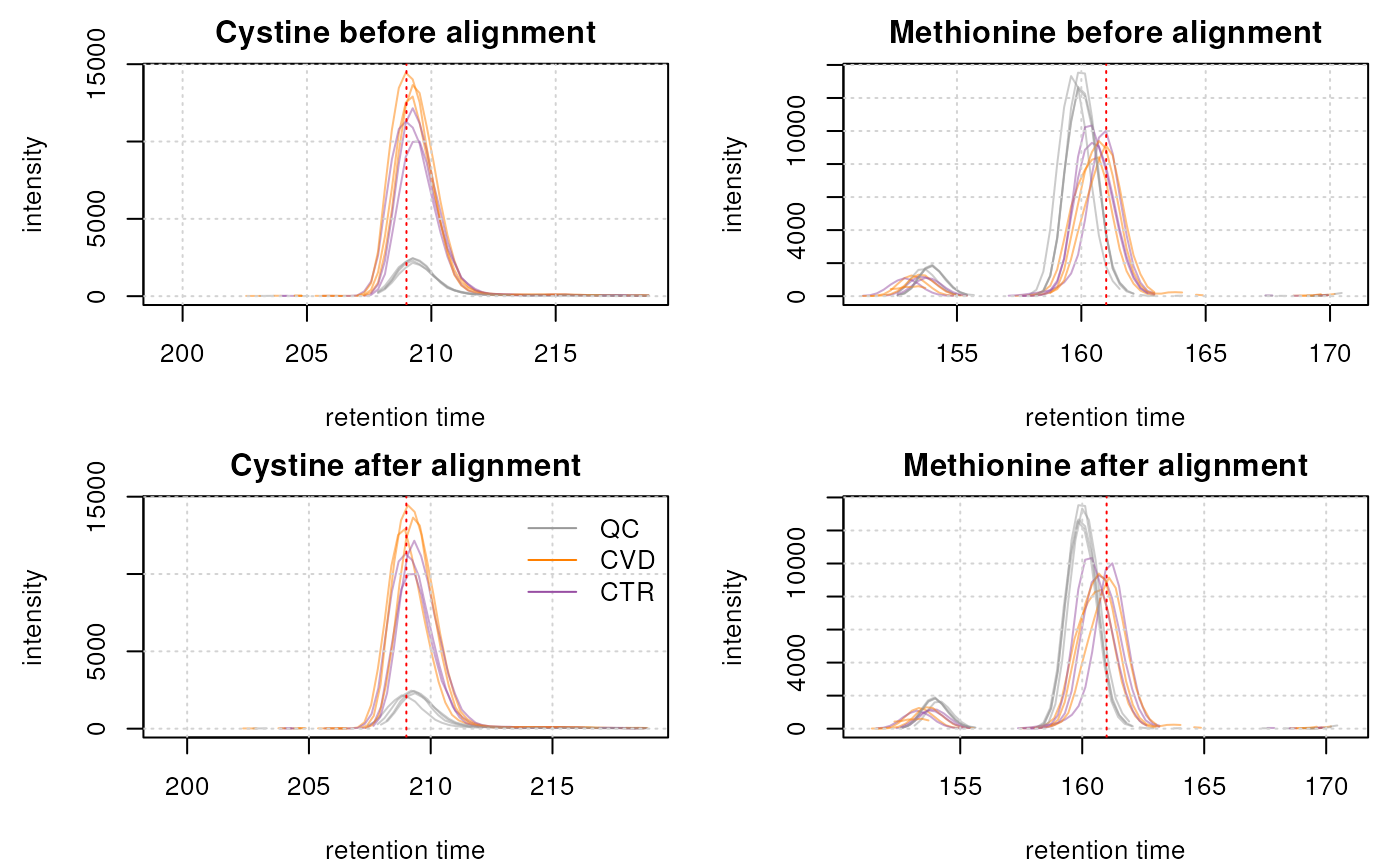
The non-endogenous cystine ion was already well aligned so the difference is minimal. The methionine ion, however, shows an improvement in alignment.
In addition to a visual inspection of the results, we also evaluate
the impact of the alignment by comparing the variance in retention times
for internal standards before and after alignment. To this end, we first
need to identify chromatographic peaks in each sample with an
m/z and retention time close to the expected values for each
internal standard. For this we use the matchValues()
function from the MetaboAnnotation
package (Rainer et al. 2022) using the
MzRtParam method to identify all chromatographic peaks with
similar m/z (+/- 50 ppm) and retention time (+/- 10 seconds) to
the internal standard’s values. With parameters mzColname
and rtColname we specify the column names in the query (our
IS) and target (chromatographic peaks) that contain the m/z and
retention time values on which to match entities. We perform this
matching separately for each sample. For each internal standard in every
sample, we use the filterMatches() function and the
SingleMatchParam() parameter to select the chromatographic
peak with the highest intensity.
#' Extract the matrix with all chromatographic peaks and add a column
#' with the ID of the chromatographic peak
chrom_peaks <- chromPeaks(data) |> as.data.frame()
chrom_peaks$peak_id <- rownames(chrom_peaks)
#' Define the parameters for the matching and filtering of the matches
p_1 <- MzRtParam(ppm = 50, toleranceRt = 10)
p_2 <- SingleMatchParam(duplicates = "top_ranked", column = "target_maxo",
decreasing = TRUE)
#' Iterate over samples and identify for each the chromatographic peaks
#' with similar m/z and retention time than the onse from the internal
#' standard, and extract among them the ID of the peaks with the
#' highest intensity.
intern_standard_peaks <- lapply(seq_along(data), function(i) {
tmp <- chrom_peaks[chrom_peaks[, "sample"] == i, , drop = FALSE]
mtch <- matchValues(intern_standard, tmp,
mzColname = c("mz", "mz"),
rtColname = c("RT", "rt"),
param = p_1)
mtch <- filterMatches(mtch, p_2)
mtch$target_peak_id
}) |>
do.call(what = cbind)We have now for each internal standard the ID of the chromatographic peak in each sample that most likely represents signal from that ion. We can now extract the retention times for these chromatographic peaks before and after alignment.
#' Define the index of the selected chromatographic peaks in the
#' full chromPeaks matrix
idx <- match(intern_standard_peaks, rownames(chromPeaks(data)))
#' Extract the raw retention times for these
rt_raw <- chromPeaks(data_raw)[idx, "rt"] |>
matrix(ncol = length(data_raw))
#' Extract the adjusted retention times for these
rt_adj <- chromPeaks(data)[idx, "rt"] |>
matrix(ncol = length(data_raw))We can now evaluate the impact of the alignment on the retention times of internal standards across the full data set:
list(all_raw = rowSds(rt_raw, na.rm = TRUE),
all_adj = rowSds(rt_adj, na.rm = TRUE)
) |>
vioplot(ylab = "sd(retention time)")
grid()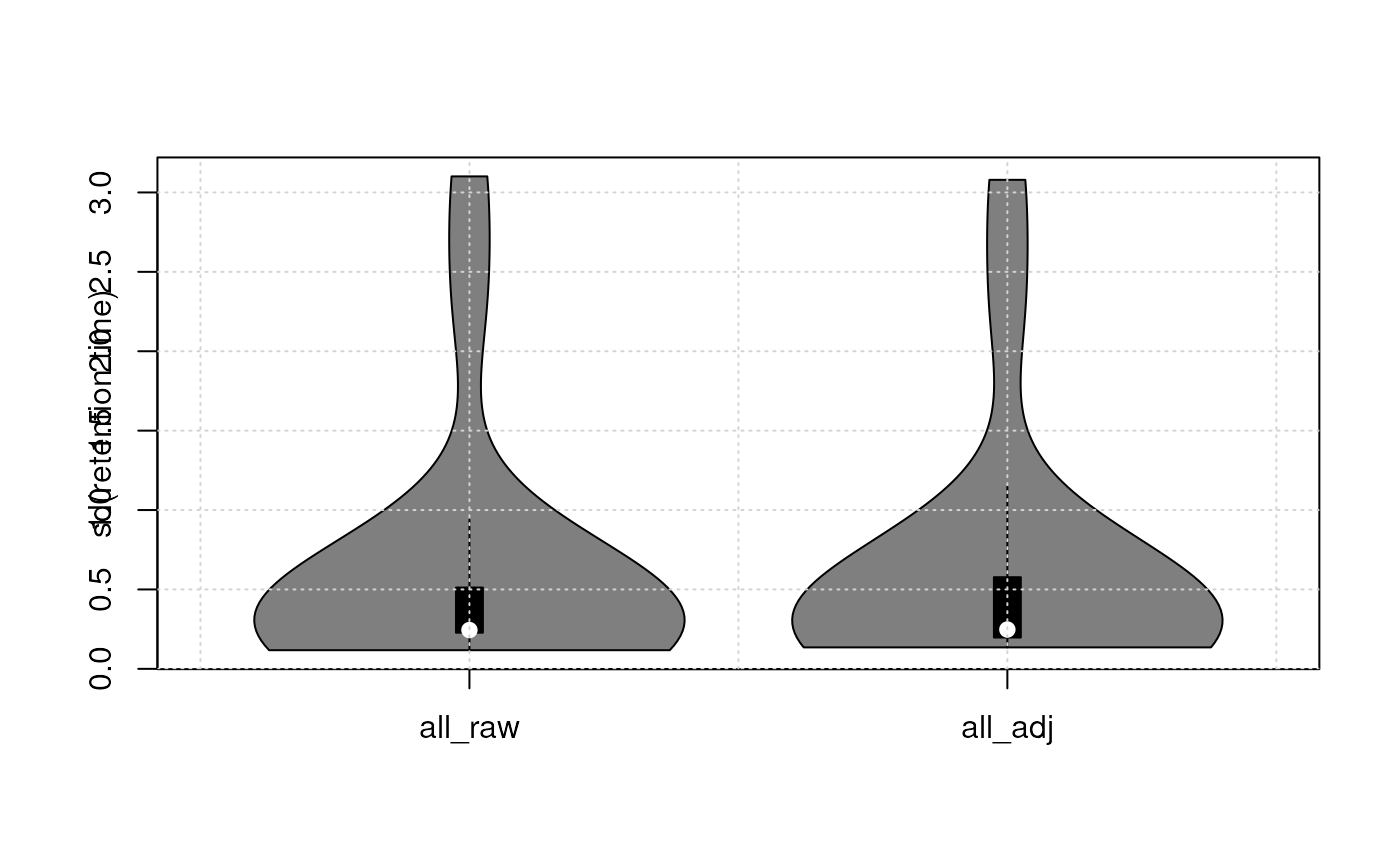
On average, the variation in retention times of internal standards across samples was slightly reduced by the alignment.[Phili: actually i don’t think we can say that at all from the plot]
Correspondence
We briefly touched on the subject of correspondence before to
determine anchor peaks for alignment. Generally, the goal of the
correspondence analysis is to identify chromatographic peaks that
originate from the same types of ions in all samples of an experiment
and to group them into LC-MS features. At this point, proper
configuration of parameter bw is crucial. Here we
illustrate how sensible choices for this parameter’s value can be made.
We use below the plotChromPeakDensity() function to
simulate a correspondence analysis with the default values for
PeakGroups on the extracted ion chromatograms of our two
selected isotope labeled ions. This plot shows the EIC in the top panel,
and the apex position of chromatographic peaks in the different samples
(y-axis), along retention time (x-axis) in the lower panel.
#' Default parameter for the grouping and apply them to the test ions BPC
param <- PeakDensityParam(sampleGroups = sampleData(data)$phenotype, bw = 30)
param## Object of class: PeakDensityParam
## Parameters:
## - sampleGroups: [1] "QC" "CVD" "CTR" "QC" "CTR" "CVD" "QC" "CTR" "CVD" "QC"
## - bw: [1] 30
## - minFraction: [1] 0.5
## - minSamples: [1] 1
## - binSize: [1] 0.25
## - maxFeatures: [1] 50
## - ppm: [1] 0
plotChromPeakDensity(
eic_cystine, param = param,
col = paste0(col_sample, "80"),
peakCol = col_sample[chromPeaks(eic_cystine)[, "sample"]],
peakBg = paste0(col_sample[chromPeaks(eic_cystine)[, "sample"]], 20),
peakPch = 16)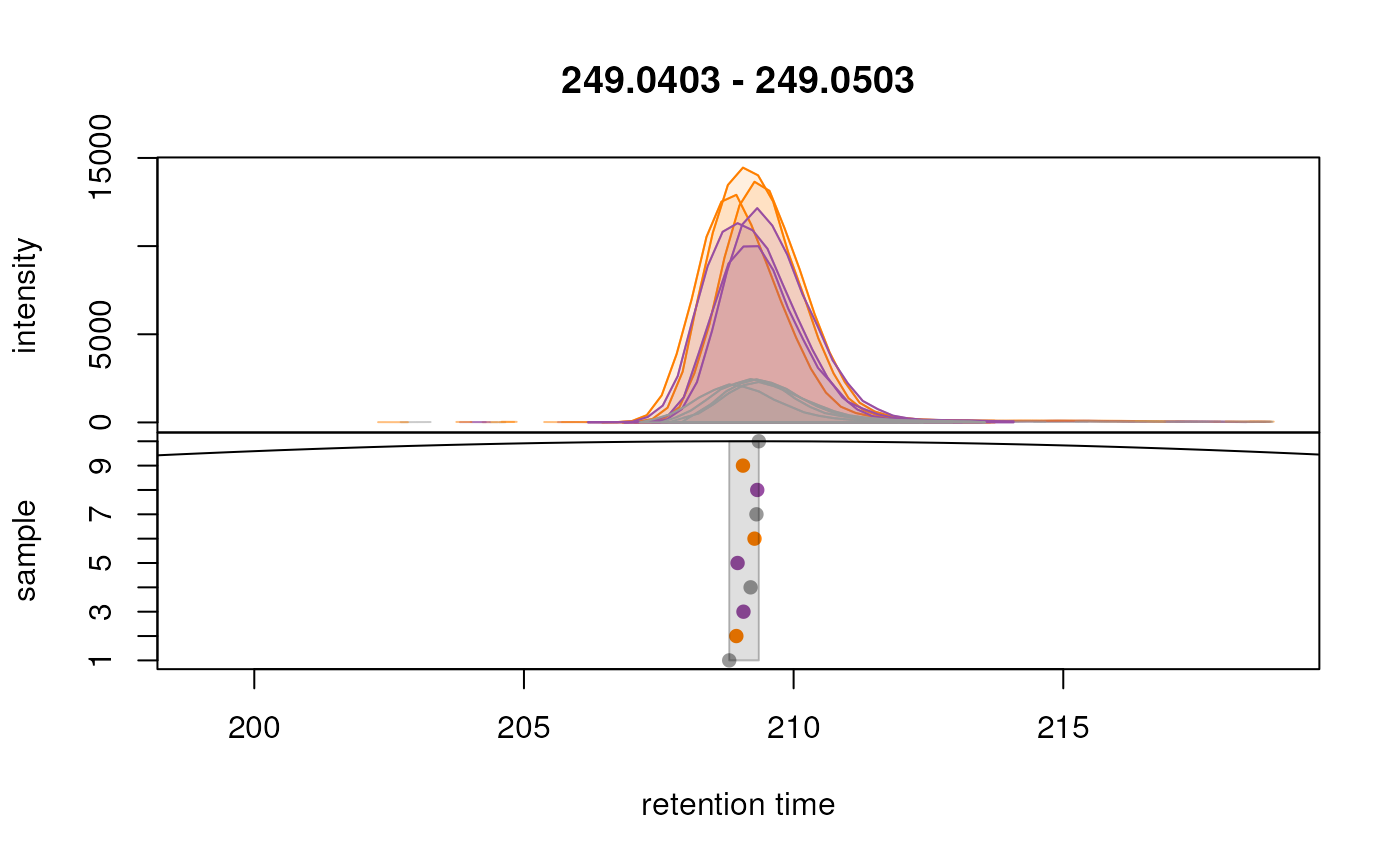
plotChromPeakDensity(eic_met, param = param,
col = paste0(col_sample, "80"),
peakCol = col_sample[chromPeaks(eic_met)[, "sample"]],
peakBg = paste0(col_sample[chromPeaks(eic_met)[, "sample"]], 20),
peakPch = 16)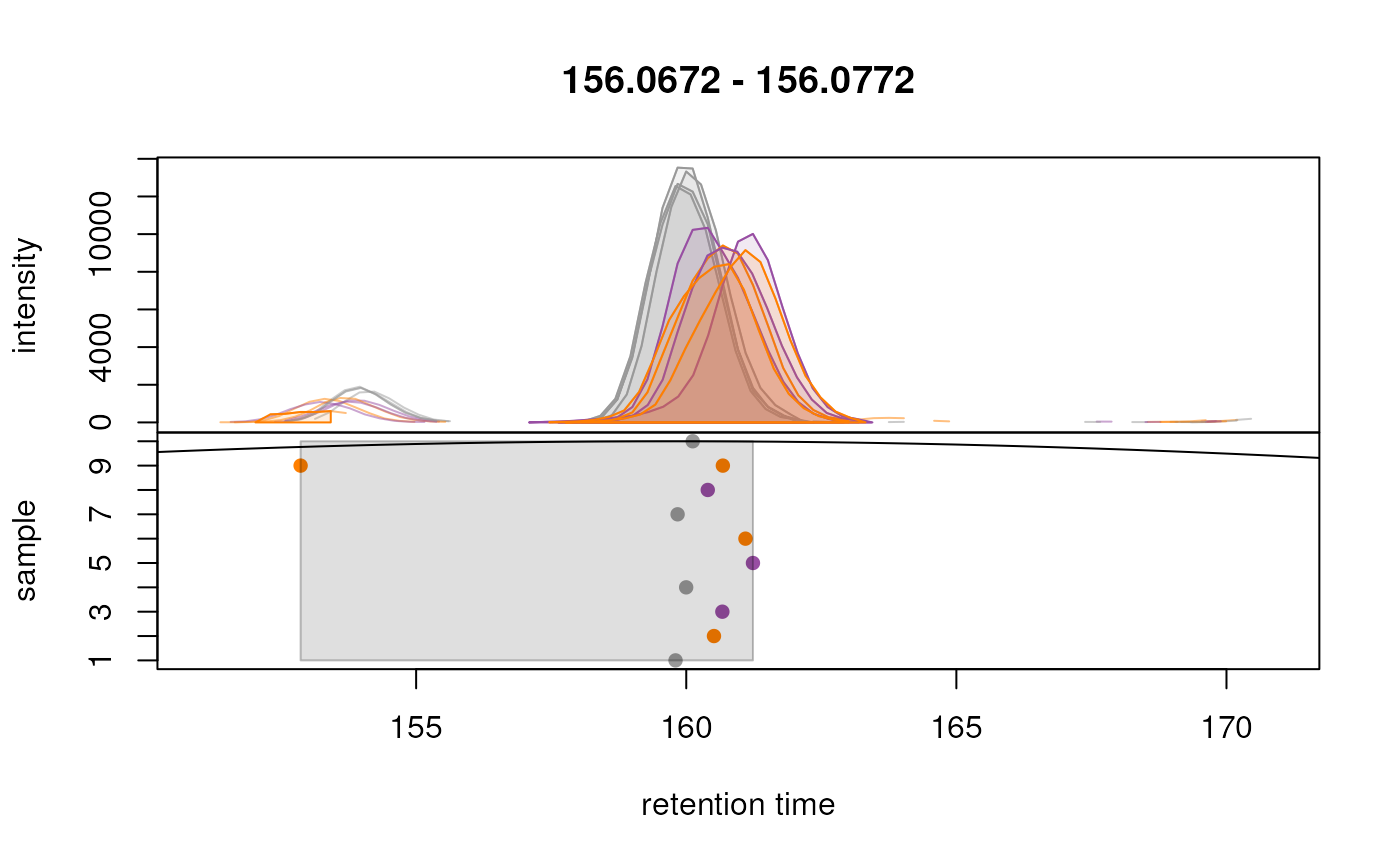
Grouping of peaks depends on the smoothness of the previousl
mentionned density curve that can be configured with the parameter
bw. As seen above, the smoothness is too high to properly
group our features. When looking at the default parameters, we can
observe that indeed, the bw parameter is set to
bw = 30, which is too high for modern UHPLC-MS setups. We
reduce the value of this parameter to 1.8 and evaluate its impact.
#' Updating parameters
param <- PeakDensityParam(sampleGroups = sampleData(data)$phenotype, bw = 1.8)
plotChromPeakDensity(
eic_cystine, param = param,
col = paste0(col_sample, "80"),
peakCol = col_sample[chromPeaks(eic_cystine)[, "sample"]],
peakBg = paste0(col_sample[chromPeaks(eic_cystine)[, "sample"]], 20),
peakPch = 16)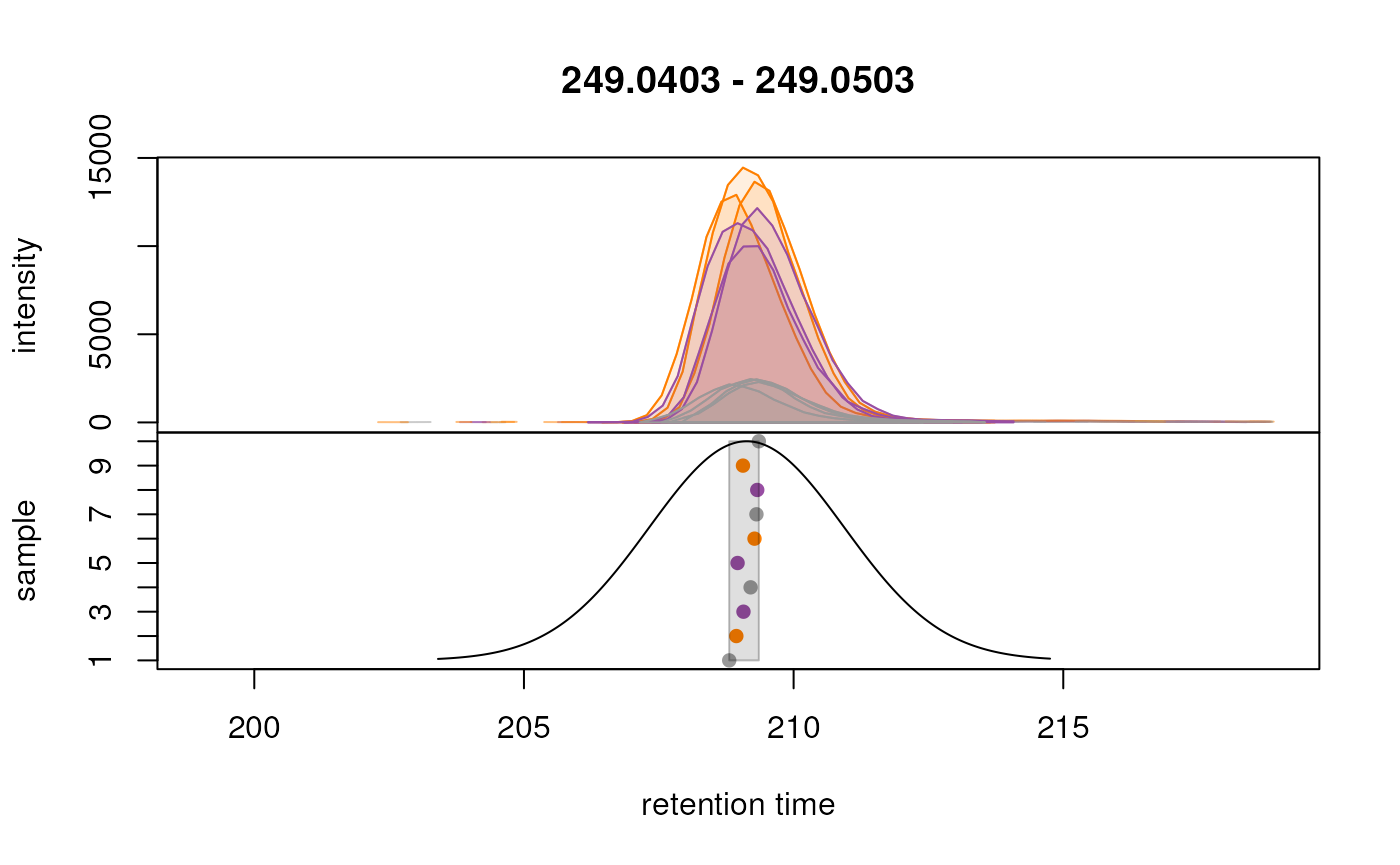
plotChromPeakDensity(eic_met, param = param,
col = paste0(col_sample, "80"),
peakCol = col_sample[chromPeaks(eic_met)[, "sample"]],
peakBg = paste0(col_sample[chromPeaks(eic_met)[, "sample"]], 20),
peakPch = 16)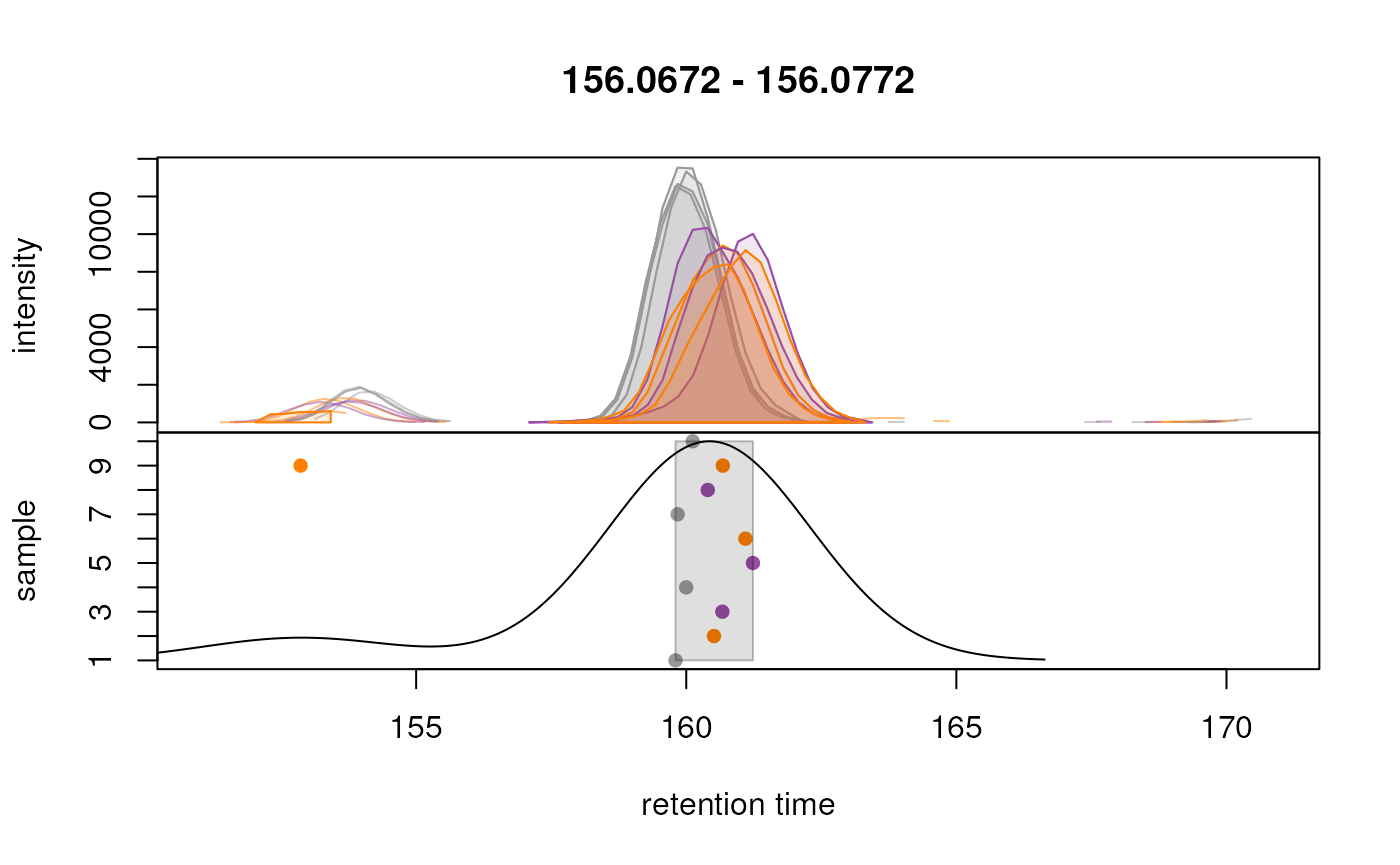
We can observe that the peaks are now grouped more accurately into a single feature for each test ion. The other important parameters optimized here are:
binsize: Our data was generated on a high resolution MS instrument, thus we select a low value for this paramete.ppm: For TOF instruments, it is suggested to use a value forppmlarger than 0 to accommodate the higher measurement error of the instrument for larger m/z values.minFraction: We set it tominFraction = 0.75, hence defining features only if a chromatographic peak was identified in at least 75% of the samples of one of the sample groups.sampleGroups: We use the information available in oursampleData’s"phenotype"column.
#' Define the settings for the param
param <- PeakDensityParam(sampleGroups = sampleData(data)$phenotype,
minFraction = 0.75, binSize = 0.01, ppm = 10,
bw = 1.8)
#' Apply to whole data
data <- groupChromPeaks(data, param = param)After correspondence analysis it is suggested to evaluate the results
again for selected EICs. Below we extract signal for an m/z
similar to that of the isotope labeled methionine for a larger retention
time range. Importantly, to show the actual correspondence results, we
set simulate = FALSE for the
plotChromPeakDensity() function.
#' Extract chromatogram for an m/z similar to the one of the labeled methionine
chr_test <- chromatogram(data,
mz = as.matrix(intern_standard["methionine_13C_15N",
c("mzmin", "mzmax")]),
rt = c(145, 200),
aggregationFun = "max")
plotChromPeakDensity(
chr_test, simulate = FALSE,
col = paste0(col_sample, "80"),
peakCol = col_sample[chromPeaks(chr_test)[, "sample"]],
peakBg = paste0(col_sample[chromPeaks(chr_test)[, "sample"]], 20),
peakPch = 16)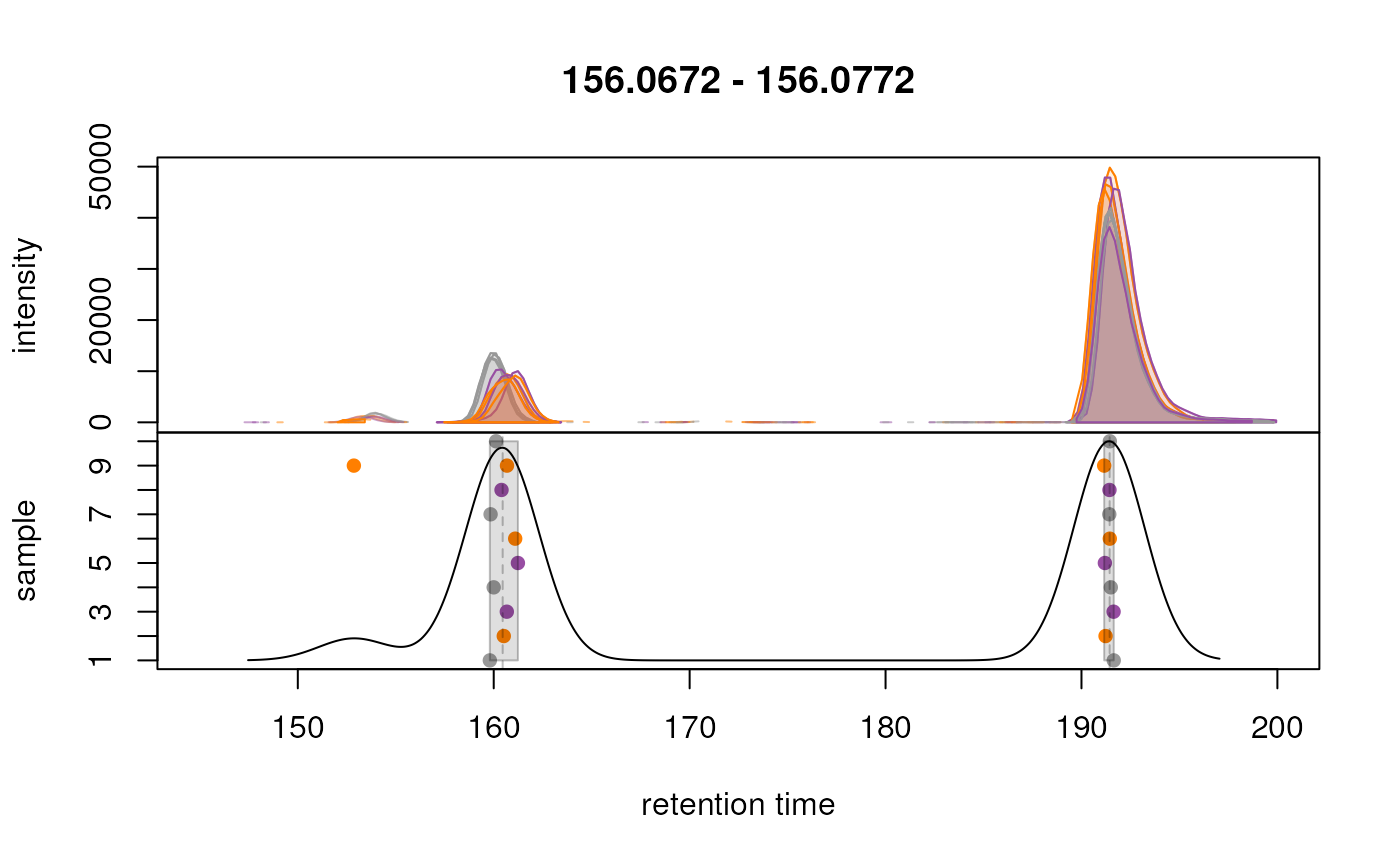
As hoped, signal from the two different ions are now grouped into separate features. Generally, correspondence results should be evaluated on more such extracted chromatograms.
Another interesting information to look a the the distribution of these features along the retention time axis.
# Bin features per RT slices
vc <- featureDefinitions(data)$rtmed
breaks <- seq(0, max(vc, na.rm = TRUE) + 1, length.out = 15) |>
round(0)
cuts <- cut(vc, breaks = breaks, include.lowest = TRUE)
table(cuts) |>
pandoc.table(
style = "rmarkdown",
caption = "Table 5.Distribution of features along the retention time axis (in seconds.")| [0,17] | (17,34] | (34,51] | (51,68] | (68,86] | (86,103] | (103,120] |
|---|---|---|---|---|---|---|
| 15 | 2608 | 649 | 24 | 132 | 11 | 15 |
| (120,137] | (137,154] | (154,171] | (171,188] | (188,205] | (205,222] |
|---|---|---|---|---|---|
| 105 | 1441 | 2202 | 410 | 869 | 554 |
| (222,240] |
|---|
| 33 |
The results from the correspondence analysis are now stored, along
with the results from the other preprocessing steps, within our
XcmsExperiment result object. The correspondence results,
i.e., the definition of the LC-MS features, can be extracted using the
featureDefinitions() function.
#' Definition of the features
featureDefinitions(data) |>
head()## mzmed mzmin mzmax rtmed rtmin rtmax npeaks CTR CVD QC
## FT0001 50.98979 50.98949 50.99038 203.6001 203.1181 204.2331 8 1 3 4
## FT0002 51.05904 51.05880 51.05941 191.1675 190.8787 191.5050 9 2 3 4
## FT0003 51.98657 51.98631 51.98699 203.1467 202.6406 203.6710 7 0 3 4
## FT0004 53.02036 53.02009 53.02043 203.2343 202.5652 204.0901 10 3 3 4
## FT0005 53.52080 53.52051 53.52102 203.1936 202.8490 204.0901 10 3 3 4
## FT0006 54.01007 54.00988 54.01015 159.2816 158.8499 159.4484 6 1 3 2
## peakidx ms_level
## FT0001 7702, 16.... 1
## FT0002 7176, 16.... 1
## FT0003 7680, 17.... 1
## FT0004 7763, 17.... 1
## FT0005 8353, 17.... 1
## FT0006 5800, 15.... 1This data frame provides the average m/z and retention time
(in columns "mzmed" and "rtmed") that
characterize a LC-MS feature. Column, "peakidx" contains
the indices of all chromatographic peaks that were assigned to that
feature. The actual abundances for these features, which represent also
the final preprocessing results, can be extracted with the
featureValues() function:
#' Extract feature abundances
featureValues(data, method = "sum") |>
head()## MS_QC_POOL_1_POS.mzML MS_A_POS.mzML MS_B_POS.mzML MS_QC_POOL_2_POS.mzML
## FT0001 421.6162 689.2422 NA 481.7436
## FT0002 710.8078 875.9192 NA 693.6997
## FT0003 445.5711 613.4410 NA 497.8866
## FT0004 16994.5260 24605.7340 19766.707 17808.0933
## FT0005 3284.2664 4526.0531 3521.822 3379.8909
## FT0006 10681.7476 10009.6602 NA 10800.5449
## MS_C_POS.mzML MS_D_POS.mzML MS_QC_POOL_3_POS.mzML MS_E_POS.mzML
## FT0001 NA 635.2732 439.6086 570.5849
## FT0002 781.2416 648.4344 700.9716 1054.0207
## FT0003 NA 634.9370 449.0933 NA
## FT0004 22780.6683 22873.1061 16965.7762 23432.1252
## FT0005 4396.0762 4317.7734 3270.5290 4533.8667
## FT0006 NA 7296.4262 NA 9236.9799
## MS_F_POS.mzML MS_QC_POOL_4_POS.mzML
## FT0001 579.9360 437.0340
## FT0002 534.4577 711.0361
## FT0003 461.0465 232.1075
## FT0004 22198.4607 16796.4497
## FT0005 4161.0132 3142.2268
## FT0006 6817.8785 NAWe can note that some features (e.g. F0003 and F0006) have missing values in some samples. This is expected to a certain degree as not in all samples features, respectively their ions, need to be present. We will address this in the next section.
Gap filling
The previously observed missing values (NA) could be attributed to various reasons. Although they might represent a genuinely missing value, indicating that an ion (feature) was truly not present in a particular sample, they could also be a result of a failure in the preceding chromatographic peak detection step. It is crucial to be able to recover missing values of the latter category as much as possible to reduce the eventual need for data imputation. We next examine how prevalent missing values are in our present dataset:
#' Percentage of missing values
sum(is.na(featureValues(data))) /
length(featureValues(data)) * 100## [1] 26.41597We can observe a substantial number of missing values values in our dataset. Let’s therefore delve into the process of gap-filling. We first evaluate some example features for which a chromatographic peak was only detected in some samples:
ftidx <- which(is.na(rowSums(featureValues(data))))
fts <- rownames(featureDefinitions(data))[ftidx]
farea <- featureArea(data, features = fts[1:2])
chromatogram(data[c(2, 3)],
rt = farea[, c("rtmin", "rtmax")],
mz = farea[, c("mzmin", "mzmax")]) |>
plot(col = c("red", "blue"), lwd = 2)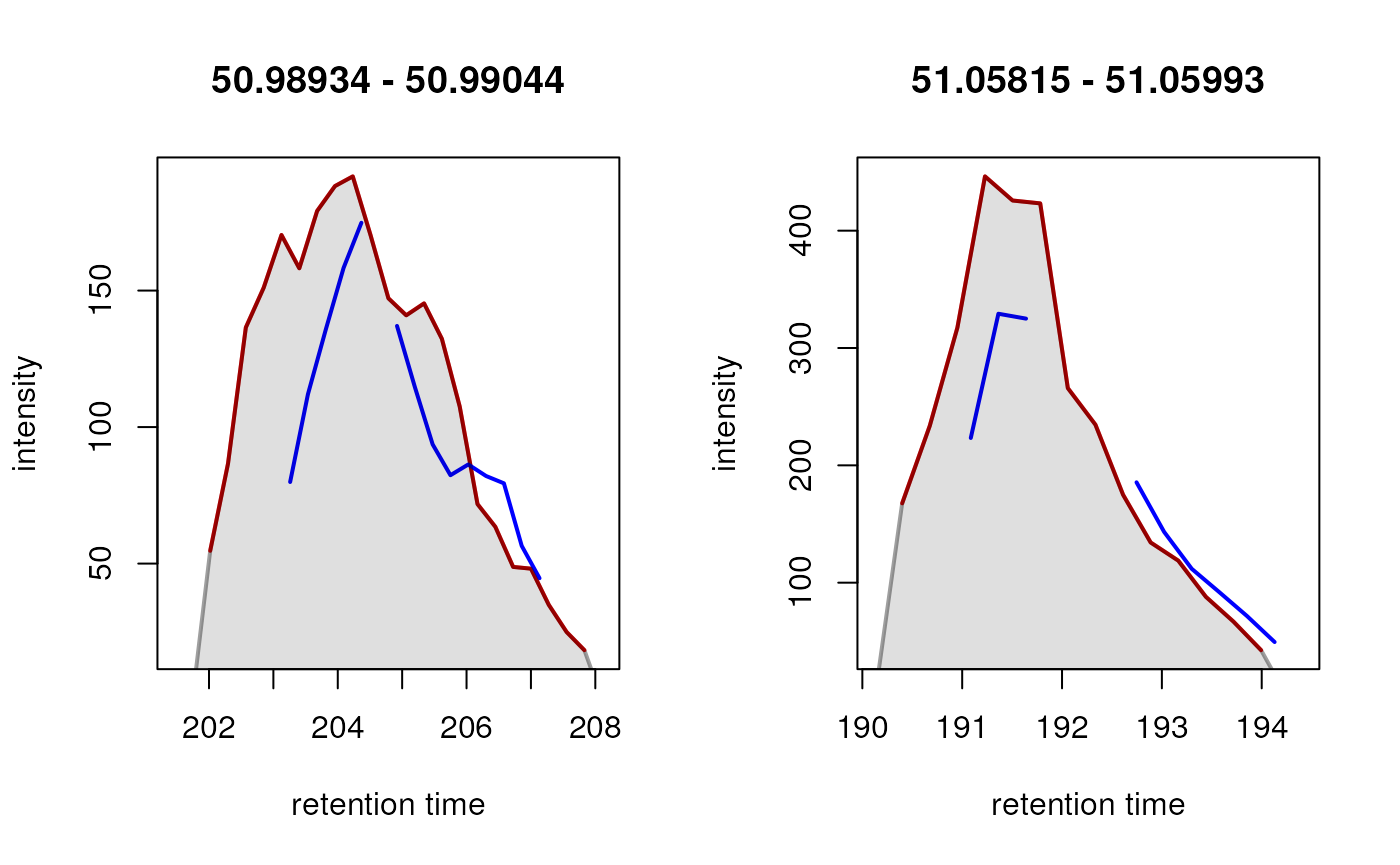
In both instances, a chromatographic peak was only identified in one
of the two selected samples (red line), hence a missing value is
reported for this feature in those particular samples (blue line).
However, in both cases, signal was measured in both samples, thus,
reporting a missing value would not be correct in this example. The
signal for this feature is very low, which is the most likely reason
peak detection failed. To rescue signal in such cases, the
fillChromPeaks() function can be used with the
ChromPeakAreaParam approach. This method defines an
m/z-retention time area for each feature based on detected
peaks, where the signal for the respective ion is expected. It then
integrates all intensities within this area in samples that have missing
values for that feature. This is then reported as feature abundance.
Below we apply this method using the default parameters.
#' Fill in the missing values in the whole dataset
data <- fillChromPeaks(data, param = ChromPeakAreaParam(), chunkSize = 5)
#' Percentage of missing values after gap-filling
sum(is.na(featureValues(data))) /
length(featureValues(data)) * 100## [1] 5.155492With fillChromPeaks() we could thus rescue most of the
missing data in the data set. Note that, even if in a sample no ion
would be present, in the worst case noise would be integrated, which is
expected to be much lower than actual chromatographic peak signal. Let’s
look at our previously missing values again:
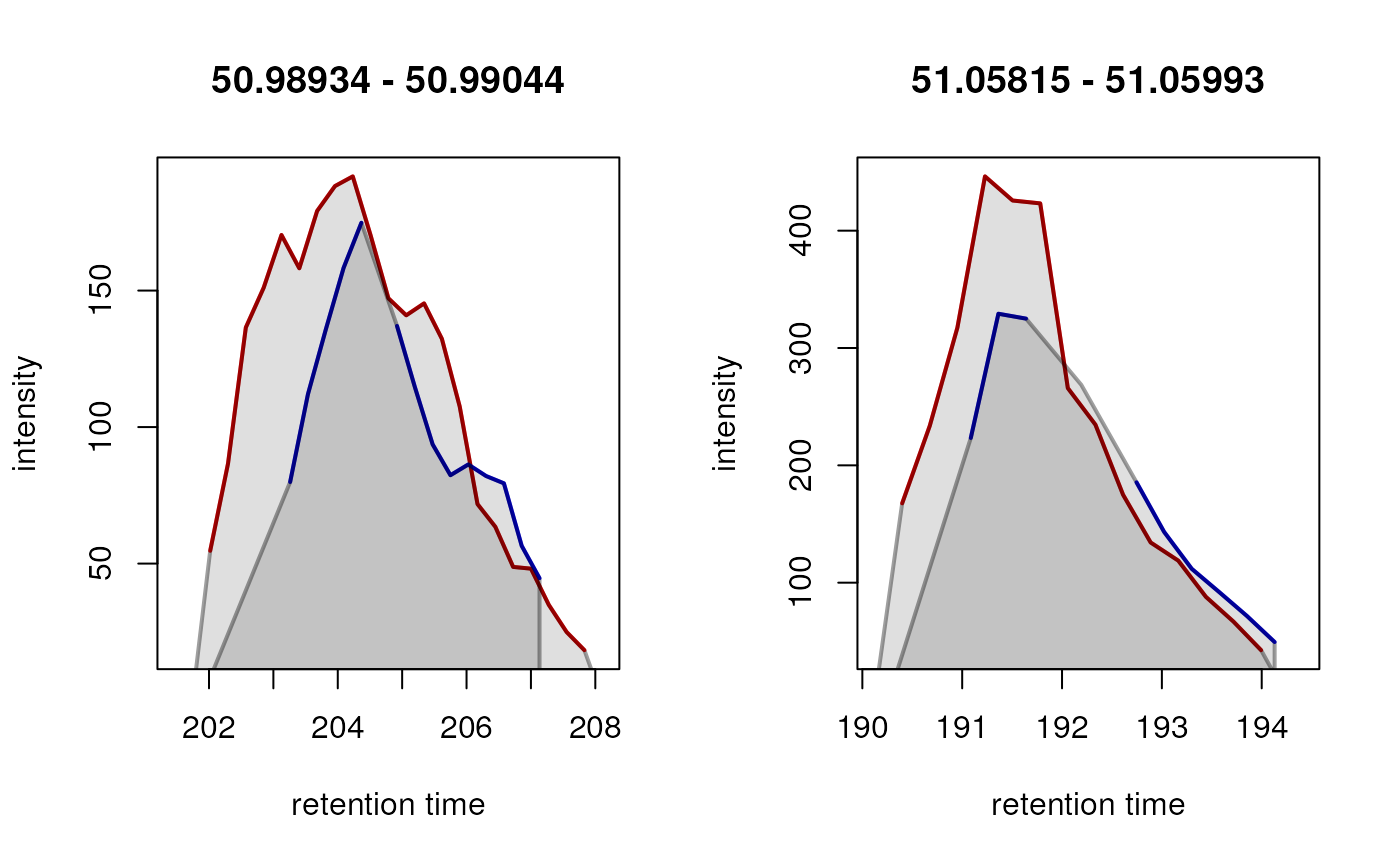
After gap-filling, also in the blue colored sample a chromatographic peak is present and its peak area would be reported as feature abundance for that sample.
To further assess the effectiveness of the gap-filling method for rescuing signals, we can also plot the average signal of features with at least one missing value against the average of filled-in signal. It is advisable to perform this analysis on repeatedly measured samples; in this case, our QC samples will be used.
For this, we extract:
Feature values from detected chromatographic peaks by setting
filled = FALSEin thefeaturesValues()call.The filled-in signal by first extracting both detected and gap-filled abundances and then replace the values for detected chromatographic peaks with
NA.
Then, we calculate the row averages of both of these matrices and plot them against each other.
#' Get only detected signal in QC samples
vals_detect <- featureValues(data, filled = FALSE)[, QC_samples]
#' Get detected and filled-in signal
vals_filled <- featureValues(data)[, QC_samples]
#' Replace detected signal with NA
vals_filled[!is.na(vals_detect)] <- NA
#' Identify features with at least one filled peak
has_filled <- is.na(rowSums(vals_detect))
#' Calculate row averages for features with missing values
avg_detect <- rowMeans(vals_detect[has_filled, ], na.rm = TRUE)
avg_filled <- rowMeans(vals_filled[has_filled, ], na.rm = TRUE)
#' Plot the values against each other (in log2 scale)
plot(log2(avg_detect), log2(avg_filled),
xlim = range(log2(c(avg_detect, avg_filled)), na.rm = TRUE),
ylim = range(log2(c(avg_detect, avg_filled)), na.rm = TRUE),
pch = 21, bg = "#00000020", col = "#00000080")
grid()
abline(0, 1)
The detected (x-axis) and gap-filled (y-axis) values for QC samples are highly correlated. Especially for higher abundances, the agreement is very high, while for low intensities, as can be expected, differences are higher and trending to below the correlation line. Below we, in addition, fit a linear regression line to the data and summarize its results
##
## Call:
## lm(formula = log2(avg_filled) ~ log2(avg_detect))
##
## Residuals:
## Min 1Q Median 3Q Max
## -6.8176 -0.3807 0.1725 0.5492 6.7504
##
## Coefficients:
## Estimate Std. Error t value Pr(>|t|)
## (Intercept) -1.62359 0.11545 -14.06 <2e-16 ***
## log2(avg_detect) 1.11763 0.01259 88.75 <2e-16 ***
## ---
## Signif. codes: 0 '***' 0.001 '**' 0.01 '*' 0.05 '.' 0.1 ' ' 1
##
## Residual standard error: 0.9366 on 2846 degrees of freedom
## (846 observations deleted due to missingness)
## Multiple R-squared: 0.7346, Adjusted R-squared: 0.7345
## F-statistic: 7877 on 1 and 2846 DF, p-value: < 2.2e-16The linear regression line has a slope of 1.12 and an intercept of -1.62. This indicates that the filled-in signal is on average 1.12 times higher than the detected signal.
Preprocessing results
The final results of the LC-MS data preprocessing are stored within
the XcmsExperiment object. This includes the identified
chromatographic peaks, the alignment results, as well as the
correspondence results. In addition, to guarantee reproducibility, this
result object keeps track of all performed processing steps, including
the individual parameter objects used to configure these. The
processHistory() function returns a list of the various
applied processing steps in chronological order. Below, we extract the
information for the first step of the performed preprocessing.
#' Check first step of the process history
processHistory(data)[[1]]## Object of class "XProcessHistory"
## type: Peak detection
## date: Mon Sep 30 14:36:38 2024
## info:
## fileIndex: 1,2,3,4,5,6,7,8,9,10
## Parameter class: CentWaveParam
## MS level(s) 1The processParam() function could then be used to
extract the actual parameter class used to configure this processing
step.
The final result of the whole LC-MS data preprocessing is a two-dimensional matrix with abundances of the so-called LC-MS features in all samples. Note that at this stage of the analysis features are only characterized by their m/z and retention time and we don’t have yet any information which metabolite a feature could represent.
As we have seen before, such feature matrix can be extracted with the
featureValues() function and the corresponding feature
characteristics (i.e., their m/z and retention time values)
using the featureDefinitions() function. Thus, these two
arrays could be extracted from the xcms result object and
used/imported in other analysis packages for further processing. They
could for example also be exported to tab delimited text files, and used
in an external tool, or used, if also MS2 spectra were available, for
feature-based molecular networking in the GNPS analysis environment
(Nothias et al. 2020).
For further processing in R, if no reference or link to the raw MS
data is required, it is suggested to extract the xcms
preprocessing result using the quantify() function as a
SummarizedExperiment object, Bioconductor’s default
container for data from biological assays/experiments. This simplifies
integration with other Bioconductor analysis packages. The
quantify() function takes the same parameters than the
featureValues() function, thus, with the call below we
extract a SummarizedExperiment with only the detected, but
not gap-filled, feature abundances:
#' Extract results as a SummarizedExperiment
res <- quantify(data, method = "sum", filled = FALSE)
res## class: SummarizedExperiment
## dim: 9068 10
## metadata(6): '' '' ... '' ''
## assays(1): raw
## rownames(9068): FT0001 FT0002 ... FT9067 FT9068
## rowData names(11): mzmed mzmin ... QC ms_level
## colnames(10): MS_QC_POOL_1_POS.mzML MS_A_POS.mzML ... MS_F_POS.mzML
## MS_QC_POOL_4_POS.mzML
## colData names(11): sample_name derived_spectra_data_file ... phenotype
## injection_indexSample identifications from the xcms result’s
sampleData() are now available as colData()
(column, sample annotations) and the featureDefinitions()
as rowData() (row, feature annotations). The feature values
have been added as the first assay() in the
SummarizedExperiment and even the processing history is
available in the object’s metadata(). A
SummarizedExperiment supports multiple assays, all being
numeric matrices with the same dimensions. Below we thus add the
detected and gap-filled feature abundances as an additional assay to the
SummarizedExperiment.
assays(res)$raw_filled <- featureValues(data, method = "sum",
filled = TRUE )
#' Different assay in the SummarizedExperiment object
assayNames(res)## [1] "raw" "raw_filled"Feature abundances can be extracted with the assay()
function. Below we extract the first 6 lines of the detected and
gap-filled feature abundances:
## MS_QC_POOL_1_POS.mzML MS_A_POS.mzML MS_B_POS.mzML MS_QC_POOL_2_POS.mzML
## FT0001 421.6162 689.2422 411.3295 481.7436
## FT0002 710.8078 875.9192 457.5920 693.6997
## FT0003 445.5711 613.4410 277.5022 497.8866
## FT0004 16994.5260 24605.7340 19766.7069 17808.0933
## FT0005 3284.2664 4526.0531 3521.8221 3379.8909
## FT0006 10681.7476 10009.6602 9599.9701 10800.5449
## MS_C_POS.mzML MS_D_POS.mzML MS_QC_POOL_3_POS.mzML MS_E_POS.mzML
## FT0001 314.7567 635.2732 439.6086 570.5849
## FT0002 781.2416 648.4344 700.9716 1054.0207
## FT0003 425.3774 634.9370 449.0933 556.2544
## FT0004 22780.6683 22873.1061 16965.7762 23432.1252
## FT0005 4396.0762 4317.7734 3270.5290 4533.8667
## FT0006 4792.2390 7296.4262 2382.1788 9236.9799
## MS_F_POS.mzML MS_QC_POOL_4_POS.mzML
## FT0001 579.9360 437.0340
## FT0002 534.4577 711.0361
## FT0003 461.0465 232.1075
## FT0004 22198.4607 16796.4497
## FT0005 4161.0132 3142.2268
## FT0006 6817.8785 6911.5439An advantage, in addition to being a container for the full preprocessing results is also the possibility of an easy and intuitive creation of data subsets ensuring data integrity. It would for example be very easy to subset the full data to a selection of features and/or samples:
res[1:14, 3:8]## class: SummarizedExperiment
## dim: 14 6
## metadata(6): '' '' ... '' ''
## assays(2): raw raw_filled
## rownames(14): FT0001 FT0002 ... FT0013 FT0014
## rowData names(11): mzmed mzmin ... QC ms_level
## colnames(6): MS_B_POS.mzML MS_QC_POOL_2_POS.mzML ...
## MS_QC_POOL_3_POS.mzML MS_E_POS.mzML
## colData names(11): sample_name derived_spectra_data_file ... phenotype
## injection_indexThe XcmsExperiment object can also be saved for later
use using the storeResults() function. The data can be
exported in different formats, to enable easier integration with
non-R-based software. Currently, it is possible to export the data in
R-specific RData format or as (separate) plain text files.
Export to the community-developed open mzTab-M format is currently being
developed and will be supported in future. Below we export the
xcms result object with R’s default binary format for object
serialization.
Data normalization
After preprocessing, data normalization or scaling might need to be applied to remove any technical variances from the data. While simple approaches like median scaling can be implemented with a few lines of R code, more advanced normalization algorithms are available in packages such as Bioconductor’s preprocessCore. The comprehensive workflow “Notame” also propose a very interesting normalization approach adaptable and scalable to the user dataset (Klåvus et al. 2020).
Generally, for LC-MS data, bias can be categorized into three main groups(Broadhurst et al. 2018):
Variances introduced by sample collection and initial processing, which can include differences in sample amounts. This type of bias is expected to be sample-specific and affect all signals in a sample in the same way. Methods like median scaling, LOESS or quantiles normalization can adjust this bias.
Signal drifts along measurement of samples from an experiment. Reasons for such drifts can be related to aging of the instrumentation used (columns, detector), but also to changes in metabolite abundances and characteristics due to reactions and modifications, such as oxidation. These changes are expected to affect more the samples measured later in a run rather than the ones measured at the beginning. For this reason, this bias can play a major role in large experiments bias can play a major role in large experiments measured over a long time range and is usually considered to affect individual metabolites (or metabolite groups) differently. For adjustment, moving average or linear regression-based approaches can be used. The latter can for example be performed using the
adjust_lm()function from the MetaboCoreUtils package.Batch-related biases. These comprise any noise that is specific to a larger set of samples, which can be the set of samples measured in one LC-MS measurement run (i.e. from one analysis plate) or samples measured using a specific batch of reagents. This noise is assumed to affect all samples in one batch in the same way and linear modeling-based approaches can be used to adjust for this.
Unwanted variation can arise from various sources and is highly dependent on the experiment. Therefore, data normalization should be chosen carefully based on experimental design, statistical aims, and the balance of accuracy and precision achieved through the use of auxiliary information.
Sample preparation biases can be evaluated using internal standards, depending however also on when they were added to the sample mixes during sample processing. Repeated measurements of QC samples on the other hand allows to estimate and correct for LC-MS specific biases. Also, proper planning of an experiment, such as measurement of study samples in random order, can largely avoid biases introduced by most of the above mentioned sources of variance.
In this workflow we present some tools to assess data quality and evaluate need for normalization as well as options for normalization. For space reasons we are not able to provide solutions to adjust for all possible sources of variation.
Initial quality assessment
A principal component analysis (PCA) is a very helpful tool for an initial, unsupervised, visualization of the data that also provides insights into potential quality issues in the data. In order to apply a PCA to the measured feature abundances, we need however to impute (the still present) missing values. We assume that most of these missing values (after the gap-filling step) represent signal which is below detection limit. In such cases, missing values can be replaced with random values sampled from a uniform distribution, ranging from half of the smallest measured value to the smallest measured value for a specific feature. The uniform distribution is defined with two parameters (minimum and maximum) and all values between them have an equal probability of being selected.
Below we impute missing values with this approach and add the resulting data matrix as a new assay to our result object.
#' Load preprocessing results
## load("SumExp.RData")
## loadResults(RDataParam("data.RData"))
#' Impute missing values using an uniform distribution
na_unidis <- function(z) {
na <- is.na(z)
if (any(na)) {
min = min(z, na.rm = TRUE)
z[na] <- runif(sum(na), min = min/2, max = min)
}
z
}
#' Row-wise impute missing values and add the data as a new assay
tmp <- apply(assay(res, "raw_filled"), MARGIN = 1, na_unidis)
assays(res)$raw_filled_imputed <- t(tmp)Principal Component Analysis
PCA is a powerful tool for detecting biases in data. As a dimensionality reduction technique, it enables visualization of data in a lower-dimensional space. In the context of LC-MS data, PCA can be used to identify overall biases in batch, sample, injection index, etc. However, it is important to note that PCA is a linear method and may not be able to detect all biases in the data.
Before plotting the PCA, we apply a log2 transform, center and scale the data. The log2 transformation is applied to stabilize the variance while centering to remove dependency on absolute abundances.
#' Log2 transform and scale data
vals <- assay(res, "raw_filled_imputed") |>
log2() |>
t() |>
scale(center = TRUE, scale = TRUE)
#' Perform the PCA
pca_res <- prcomp(vals, scale = FALSE, center = FALSE)
#' Plot the results
vals_st <- cbind(vals, phenotype = res$phenotype)
pca_12 <- autoplot(pca_res, data = vals_st , colour = 'phenotype', scale = 0) +
scale_color_manual(values = col_phenotype)
pca_34 <- autoplot(pca_res, data = vals_st, colour = 'phenotype',
x = 3, y = 4, scale = 0) +
scale_color_manual(values = col_phenotype)
grid.arrange(pca_12, pca_34, ncol = 2)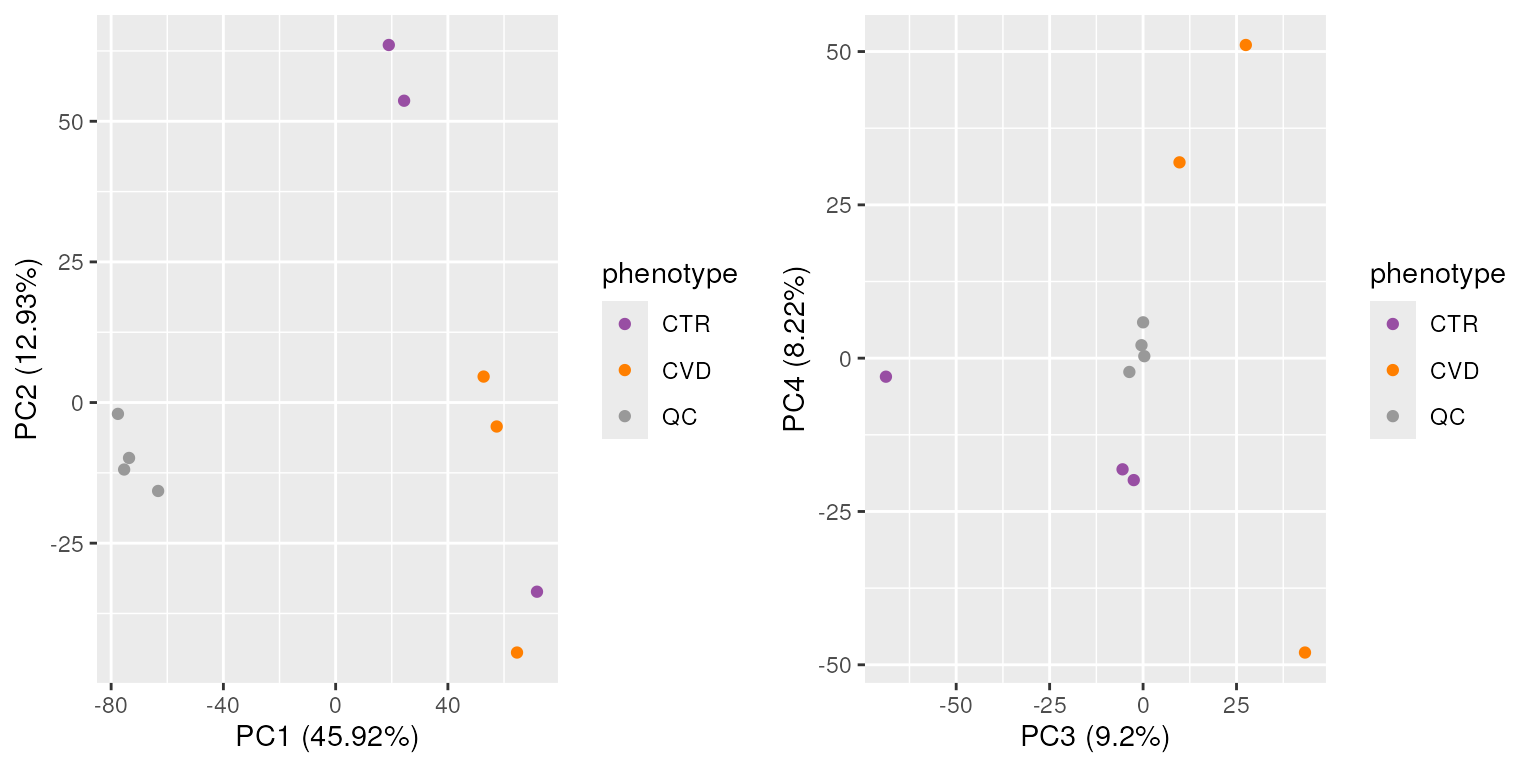
The PCA above shows a clear separation of the study samples (plasma) from the QC samples (serum) on the first principal component (PC1). The separation based on phenotype is visible on the third principal component (PC3).
In some cases, it can be a better option to remove the imputed values and evaluate the PCA again. This is especially true if the imputed values are replacing a large proportion of the data.
Intensity evaluation
Global differences in feature abundances between samples (e.g. due to sample-specific biases) can be evaluated by plotting the distribution of the log2 transformed feature abundances using boxplots of violin plots. Below we show the number of detected chromatographic peaks per sample and the distribution of the log2 transformed feature abundances.
layout(mat = matrix(1:3, ncol = 1), height = c(0.2, 0.2, 0.8))
par(mar = c(0.2, 4.5, 0.2, 3))
barplot(apply(assay(res, "raw"), MARGIN = 2, function(x) sum(!is.na(x))),
col = paste0(col_sample, 80), border = col_sample,
ylab = "# detected peaks", xaxt = "n", space = 0.012)
grid(nx = NA, ny = NULL)
barplot(apply(assay(res, "raw_filled"), MARGIN = 2, function(x) sum(!is.na(x))),
col = paste0(col_sample, 80), border = col_sample,
ylab = "# detected + filled peaks", xaxt = "n",
space = 0.012)
grid(nx = NA, ny = NULL)
vioplot(log2(assay(res, "raw_filled")), xaxt = "n",
ylab = expression(log[2]~feature~abundance),
col = paste0(col_sample, 80), border = col_sample)
points(colMedians(log2(assay(res, "raw_filled")), na.rm = TRUE), type = "b",
pch = 1)
grid(nx = NA, ny = NULL)
legend("topright", col = col_phenotype,
legend = names(col_phenotype), lty=1, lwd = 2, xpd = TRUE, ncol = 3,
cex = 0.8, bty = "n")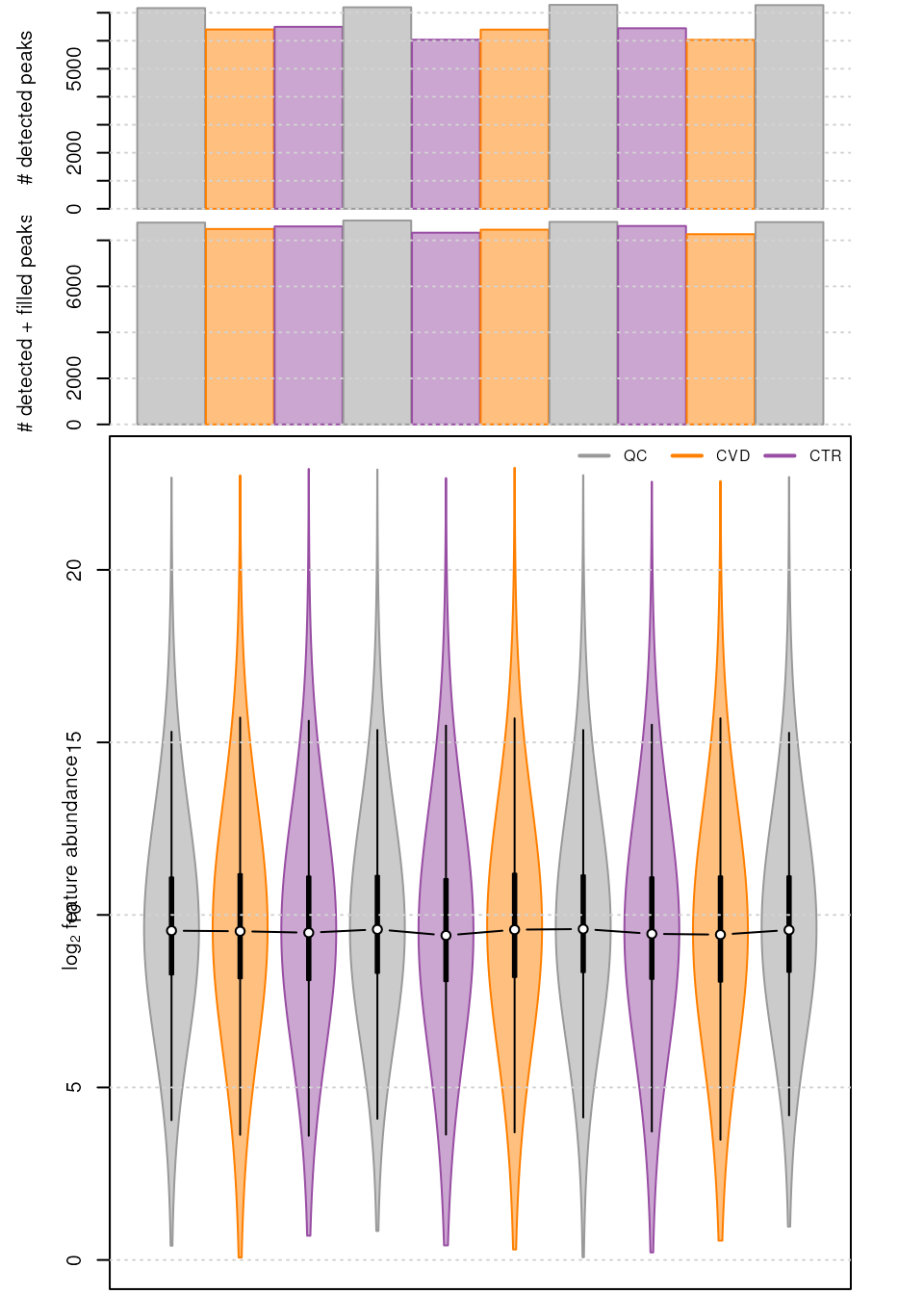
The upper part of the plot show that the gap filling steps allowed to rescue a substantial number of NAs and allowed us to have a more consistent number of feature values per sample. This consistency aligns with our asspumption that every sample should have a similar amount of features detected. Additionally we observe that, on average, the signal distribution from the individual samples is very similar.
An alternative way to evaluate differences in abundances between
samples are relative log abundance (RLA) plots (De Livera et al. 2012). An RLA value is the
abundance of a feature in a sample relative to the median abundance of
the same feature across multiple samples. We can discriminate between
within group and across group RLAs, depending whether
the abundance is compared against samples within the same sample group
or across all samples. Within group RLA plots assess the tightness of
replicates within groups and should have a median close to zero and low
variation around it. When used across groups, they allow to compare
behavior between groups. Generally, between-sample differences can be
easily spotted using RLA plots. Below we calculate and visualize within
group RLA values using the rowRla() function from the
r Biocpkg("MsCoreUtils") package defining with parameter
f the sample groups.
par(mfrow = c(1, 1), mar = c(3.5, 4.5, 2.5, 1))
boxplot(MsCoreUtils::rowRla(assay(res, "raw_filled"),
f = res$phenotype, transform = "log2"),
cex = 0.5, pch = 16,
col = paste0(col_sample, 80), ylab = "RLA",
border = col_sample, boxwex = 1,
outline = FALSE, xaxt = "n", main = "Relative log abundance",
cex.main = 1)
axis(side = 1, at = seq_len(ncol(res)), labels = colData(res)$sample_name)
grid(nx = NA, ny = NULL)
abline(h = 0, lty=3, lwd = 1, col = "black")
legend("topright", col = col_phenotype,
legend = names(col_phenotype), lty=1, lwd = 2, xpd = TRUE, ncol = 3,
cex = 0.8, bty = "n")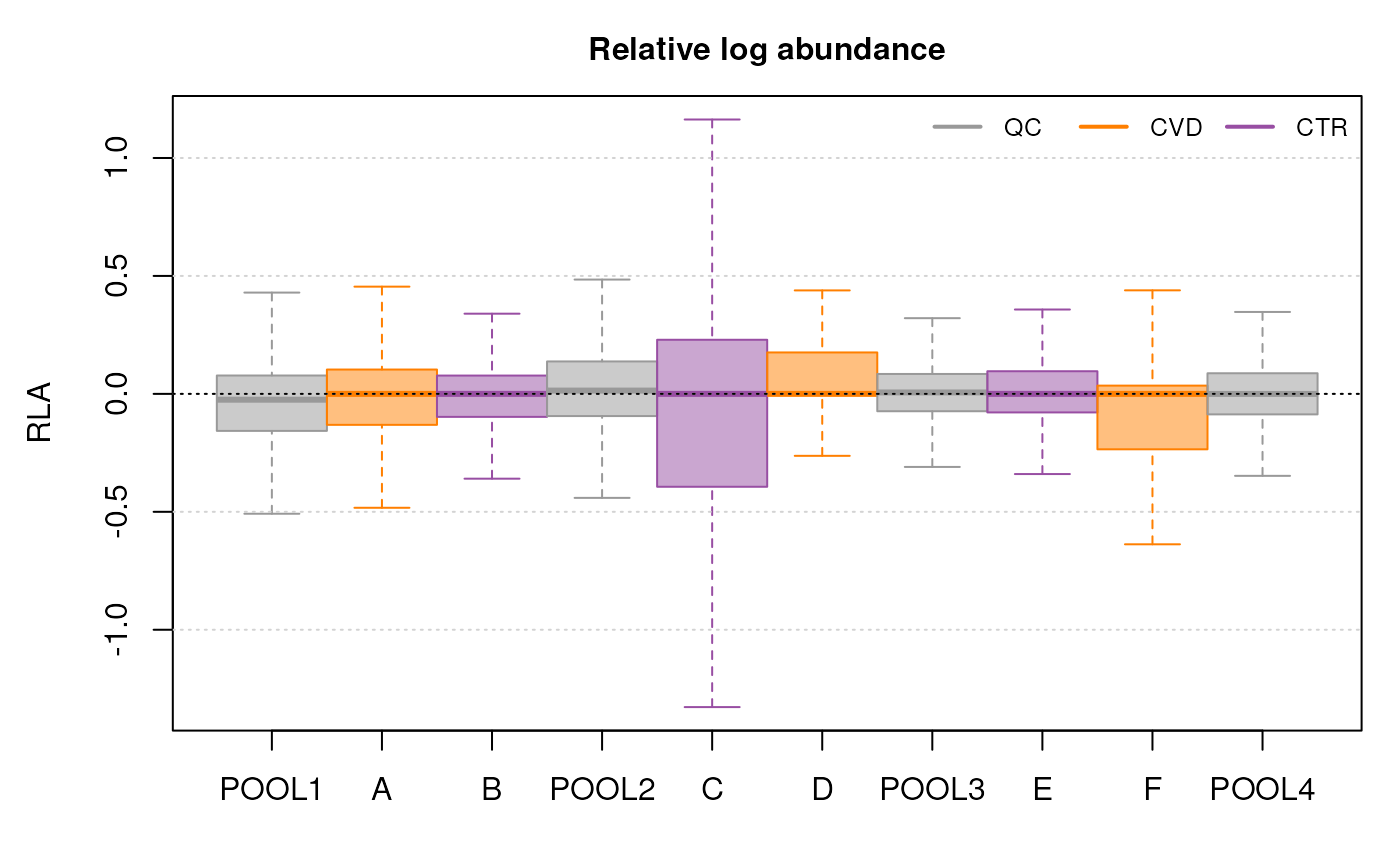
RLA plot for the raw data and filled data. Note: outliers are not drawn.
On the RLA plot above, we can observe that the medians for most samples are indeed centered around 0. Exception are two of the CVD samples. Thus, while the distribution of signals across samples is comparable, some differences seem to be present that require a between sample normalization.
Internal standards
Depending when IS are added to the sample mixes, they allow evaluation of variances introduced by the subsequent processing and analysis steps. For the present experiment, IS were added to the original plasma samples before sample extraction which included also protein and lipid removal steps. They can therefore be used to evaluate variances introduced by sample extraction and any subsequent steps, can however not be used to infer conclusions on performance or differences on the original sample collection (blood drawing, storage, plasma creation).
Here we again use the matchValues() function to identify
features representing signal from the IS. We then filter these matches
to only keep IS that match with a single feature using the
filterMatches() function in combination with
SingleMatchParam.
# Do we keep IS in normalisation ? Does not give much info... Would simplify a bit
#' Creating a column within our IS table
intern_standard$feature_id <- NA_character_
#' Identify features matching m/z and RT of internal standards.
fdef <- featureDefinitions(data)
fdef$feature_id <- rownames(fdef)
match_intern_standard <- matchValues(
query = intern_standard,
target = fdef,
mzColname = c("mz", "mzmed"),
rtColname = c("RT", "rtmed"),
param = MzRtParam(ppm = 50, toleranceRt = 10))
#' Keep only matches with a 1:1 mapping standard to feature.
param <- SingleMatchParam(duplicates = "remove", column = "score_rt",
decreasing = TRUE)
match_intern_standard <- filterMatches(match_intern_standard, param)
intern_standard$feature_id <- match_intern_standard$target_feature_id
intern_standard <- intern_standard[!is.na(intern_standard$feature_id), ]These internal standards play a crucial role in guiding the normalization process. Given the assumption that the samples were artificially spiked, we possess a known ground truth—that the abundance or intensity of the internal standard should be consistent. Any difference is expected to be due to technical differences/variance. Consequently, normalization aims to minimize variation between samples for the internal standard, reinforcing the reliability of our analyses.
Between sample normalisation
The previous RLA plot showed that the data had some biases that need to be corrected. Therefore, we will implement between-sample normalization using filled-in features. This process effectively mitigates variations influenced by technical issues, such as differences in sample preparation, processing and injection methods. In this instance, we will employ a commonly used technique known as median scaling (De Livera et al. 2012).
Median scaling
This method involves computing the median for each sample, followed by determining the median of these individual sample medians. This ensures consistent median values for each sample throughout the entire data set. Maintaining uniformity in the average total metabolite abundance across all samples is crucial for effective implementation.
This process aims to establish a shared baseline for the central tendency of metabolite abundance, mitigating the impact of sample-specific technical variations. This approach fosters a more robust and comparable analysis of the top features across the data set. The assumption is that normalizing based on the median, known for its lower sensitivity to extreme values, enhances the comparability of top features and ensures a consistent average abundance across samples.
#' Compute median and generate normalization factor
mdns <- apply(assay(res, "raw_filled"), MARGIN = 2,
median, na.rm = TRUE )
nf_mdn <- mdns / median(mdns)
#' divide dataset by median of median and create a new assay.
assays(res)$norm <- sweep(assay(res, "raw_filled"), MARGIN = 2, nf_mdn, '/')
assays(res)$norm_imputed <- sweep(assay(res, "raw_filled_imputed"), MARGIN = 2,
nf_mdn, '/')The median scaling is calculated for both imputed and non-imputed
data, with each set stored separately within the
SummarizedExperiment object. This approach facilitates
testing various normalization strategies while maintaining a record of
all processing steps undertaken, enabling easy regression to previous
stages if necessary.
Assessing overall effectiveness of the normalization approach
It is crucial to evaluate the effectiveness of the normalization process. This can be achieved by comparing the distribution of the log2 transformed feature abundances before and after normalization. Additionally, the RLA plots can be used to assess the tightness of replicates within groups and compare the behavior between groups.
Principal Component Analysis
#' Data before normalization
vals_st <- cbind(vals, phenotype = res$phenotype)
pca_raw <- autoplot(pca_res, data = vals_st,
colour = 'phenotype', scale = 0) +
scale_color_manual(values = col_phenotype)
#' Data after normalization
vals_norm <- apply(assay(res, "norm"), MARGIN = 1, na_unidis) |>
log2() |>
scale(center = TRUE, scale = TRUE)
pca_res_norm <- prcomp(vals_norm, scale = FALSE, center = FALSE)
vals_st_norm <- cbind(vals_norm, phenotype = res$phenotype)
pca_adj <- autoplot(pca_res_norm, data = vals_st_norm,
colour = 'phenotype', scale = 0) +
scale_color_manual(values = col_phenotype)
grid.arrange(pca_raw, pca_adj, ncol = 2)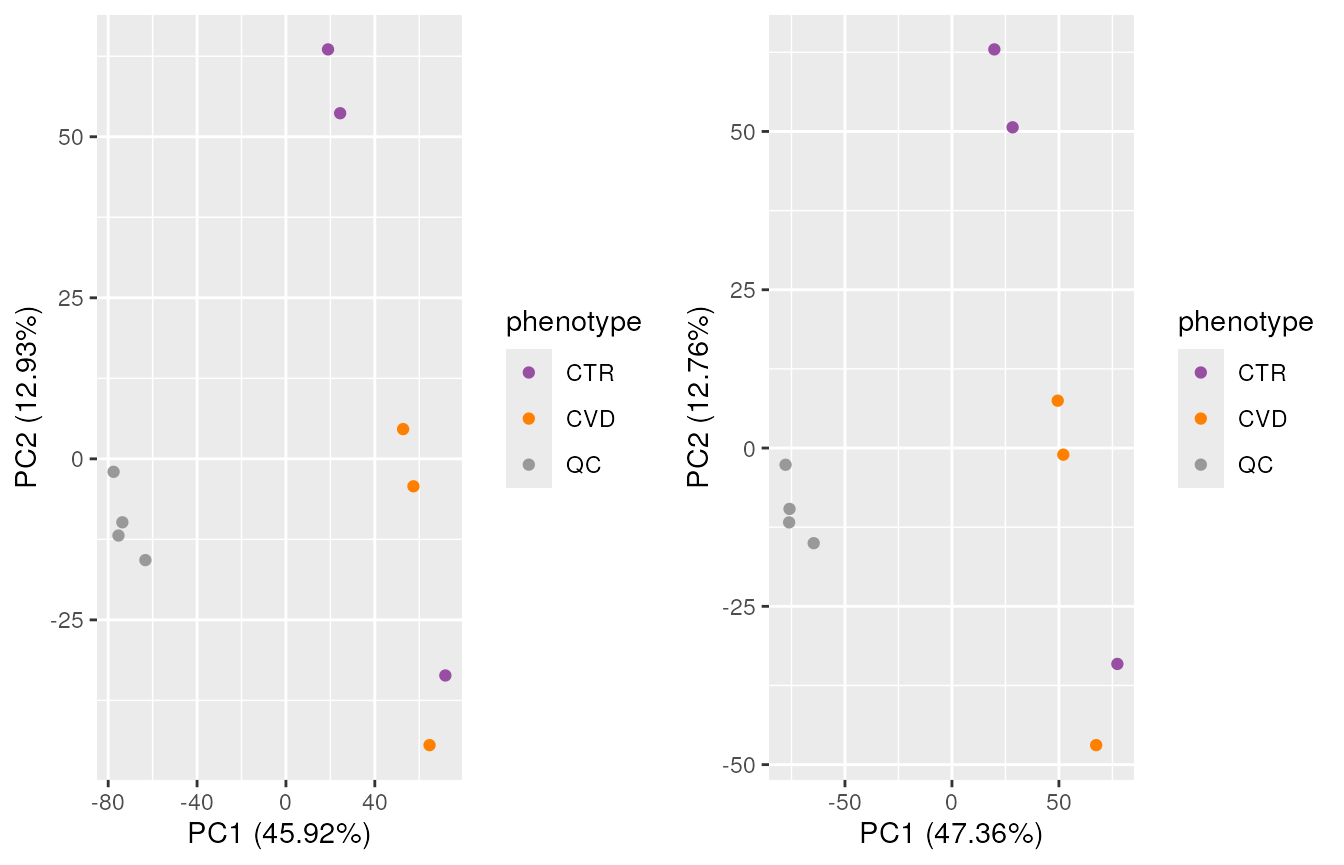
Normalization did not have a large impact on PC1 or PC2, but the separation of the study groups on PC3 seems to be better and difference between QC samples lower after normalization (see below).
pca_raw <- autoplot(pca_res, data = vals_st ,
colour = 'phenotype', x = 3, y = 4, scale = 0) +
scale_color_manual(values = col_phenotype)
pca_adj <- autoplot(pca_res_norm, data = vals_st_norm,
colour = 'phenotype', x = 3, y = 4, scale = 0) +
scale_color_manual(values = col_phenotype)
grid.arrange(pca_raw, pca_adj, ncol = 2)
The PCA plots above show that the normalization process has not changed the overall structure of the data. The separation between the study and QC samples remains the same. This is an expected results as normalization should not correct for biological variance and only technical.
Intensity evaluation
We compare the RLA plots before and after between-sample normalization to evaluate its impact on the data.
par(mfrow = c(2, 1), mar = c(3.5, 4.5, 2.5, 1))
boxplot(rowRla(assay(res, "raw_filled"), group = res$phenotype),
cex = 0.5, pch = 16, ylab = "RLA", border = col_sample,
col = paste0(col_sample, 80), cex.main = 1, outline = FALSE,
xaxt = "n", main = "Raw data", boxwex = 1)
grid(nx = NA, ny = NULL)
legend("topright", inset = c(0, -0.2), col = col_phenotype,
legend = names(col_phenotype), lty=1, lwd = 2, xpd = TRUE,
ncol = 3, cex = 0.7, bty = "n")
abline(h = 0, lty=3, lwd = 1, col = "black")
boxplot(rowRla(assay(res, "norm"), group = res$phenotype),
cex = 0.5, pch = 16, ylab = "RLA", border = col_sample,
col = paste0(col_sample, 80), boxwex = 1, outline = FALSE,
xaxt = "n", main = "Normallized data", cex.main = 1)
axis(side = 1, at = seq_len(ncol(res)), labels = res$sample_name)
grid(nx = NA, ny = NULL)
abline(h = 0, lty = 3, lwd = 1, col = "black")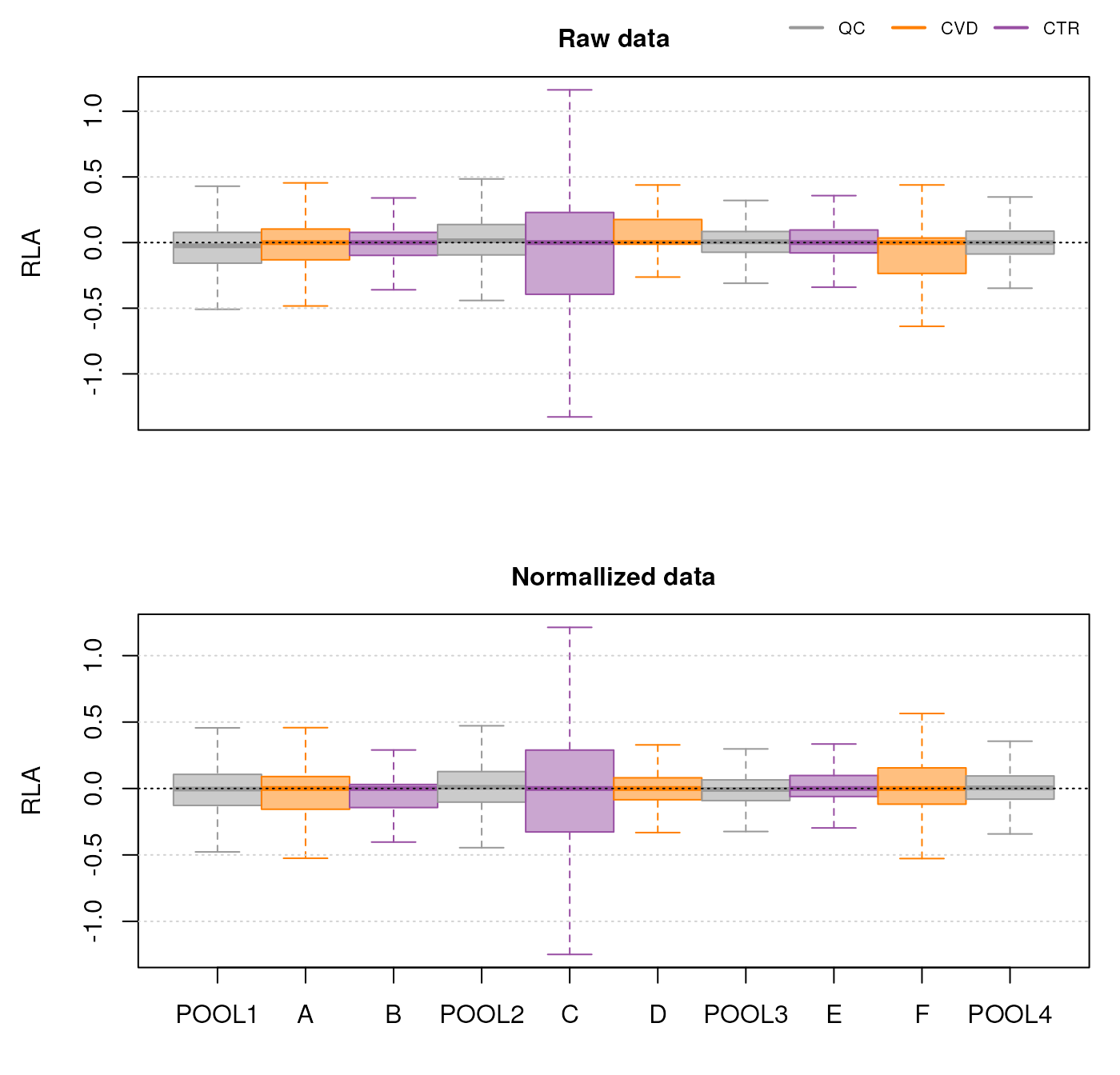
RLA plot before and after normalization. Note: outliers are not drawn.
The normalization process has effectively centered the data around the median and medians for all samples are now closer to zero.
Coefficient of variation
We next evaluate the coefficient of variation (CV, also referred to
as relative standard deviation RSD) for features across samples
from either QC or study samples. In QC samples, the CV would represent
technical noise, while in study samples it would include also expected
biological differences. Thus, normalization should reduce the CV in QC
samples, while only slightly reducing the CV in study samples. The CV is
calculated below using the rowRsd() function from the
MetaboCoreUtils
package. By setting mad = TRUE we use a more robust
calculation using the median absolute deviation instead of the standard
deviation.
index_study <- res$phenotype %in% c("CTR", "CVD")
index_QC <- res$phenotype == "QC"
sample_res <- cbind(
QC_raw = rowRsd(assay(res, "raw_filled")[, index_QC],
na.rm = TRUE, mad = TRUE),
QC_norm = rowRsd(assay(res, "norm")[, index_QC],
na.rm = TRUE, mad = TRUE),
Study_raw = rowRsd(assay(res, "raw_filled")[, index_study],
na.rm = TRUE, mad = TRUE),
Study_norm = rowRsd(assay(res, "norm")[, index_study],
na.rm = TRUE, mad = TRUE)
)
#' Summarize the values across features
res_df <- data.frame(
QC_raw = quantile(sample_res[, "QC_raw"], na.rm = TRUE),
QC_norm = quantile(sample_res[, "QC_norm"], na.rm = TRUE),
Study_raw = quantile(sample_res[, "Study_raw"], na.rm = TRUE),
Study_norm = quantile(sample_res[, "Study_norm"], na.rm = TRUE)
)
cpt <- paste0("Table 6. Distribution of CV values across samples for the raw and ",
"normalized data.")
pandoc.table(res_df, style = "rmarkdown", caption = cpt)| QC_raw | QC_norm | Study_raw | Study_norm | |
|---|---|---|---|---|
| 0% | 0 | 0 | 0 | 0 |
| 25% | 0.04557 | 0.0451 | 0.1363 | 0.1373 |
| 50% | 0.09758 | 0.09715 | 0.2635 | 0.2622 |
| 75% | 0.2003 | 0.1987 | 0.4859 | 0.4824 |
| 100% | 1.464 | 1.464 | 1.483 | 1.483 |
The table above shows the distribution of the CV for both raw and normalized data. The first column highlights the % of the data which is below a given CV value, e.g. 25% of the data has a CV equal or lower than 0.04557 at the QC_raw data. As anticipated, the CV values for the QCs, which reflect technical variance, are lower compared to those for the study samples, which include both technical and biological variance. Overall, minimal disparity exists between the raw and normalized data, which is a positive indication that the normalization process has not introduced bias into the dataset, but also reflects the little differences in average abundances between sample in the raw data.
Conclusion on normalization
The overall conclusion of the normalization process is that very
little variance was present from the beginning, normalization was
however able to center the data around the median (as shown by the RLA
plot). Given the simplicity and limited size of our example dataset, we
will conclude the normalization process at this stage. For more
intricate datasets with diverse biases, a tailored approach would be
devised. This could include also approaches to adjust for signal drifts
or batch effects. One possible option would be to use a linear-model
based approach such as can for example be applied with the
adjust_lm() function from the MetaboCoreUtils
package.
Quality control: Feature prefiltering
After normalizing our data we can now pre-filter to clean the data before performing any statistical analysis. In general, pre-filtering of samples and features is performed to remove outliers.
Below we copy the original result object to also keep the unfiltered data for later comparisons.
load("SumExp_afternorm.RData")
load("data_afternorm.RData")
#' Number of features before filtering
nrow(res)## [1] 9068
#' keep unfiltered object
res_unfilt <- resHere we will eliminate features that exhibit high variability in our dataset. Repeatedly measured QC samples typically serve as a robust basis for cleansing datasets allowing to identify features with excessively high noise. As in our data set external QC samples were used, i.e. pooled samples from a different collection and using a slightly different sample matrix, their utility for filtering is somewhat limited. For a comprehensive description and guidelines for data filtering in untargeted metabolomic studies, please refer to (Broadhurst et al. 2018).
We first restrict the data set to features for which a
chromatographic peak was detected in at least 2/3 of samples of at least
one of the study samples groups. This ensures the statistical tests
carried out later on the study samples are being performed on
reliable signal. Also, with this filter we remove features that
were mostly detected in QC samples, but not the study samples. Such
filter can be performed with filterFeatures() function from
the xcms package
with the PercentMissingFilter setting. The parameters of
this filer:
-
threshold: defines the maximal acceptable percentage of samples with missing value(s) in at least one of the sample groups defined by parameterf. -
f: a factor defining the sample groups. By replacing the"QC"sample group withNAin parameterfwe exclude the QC samples from the evaluation and consider only the study samples. Withthreshold = 40we keep only features for which a peak was detected in 2 out of the 3 samples in one of the sample groups.
To consider detected chromatographic peaks per sample, we apply the
filter on the"raw" assay of our result object, that
contains abundance values only for detected chromatographic peaks (prior
gap-filling).
#' Limit features to those with at least two detected peaks in one study group.
#' Setting the value for QC samples to NA excludes QC samples from the
#' calculation.
f <- res$phenotype
f[f == "QC"] <- NA
f <- as.factor(f)
res <- filterFeatures(res, PercentMissingFilter(f = f, threshold = 40),
assay = "raw")Following the guidelines stated above we decided to still use the QC samples for pre-filtering, on the basis that they represent similar bio-fluids to our study samples, and thus, we anticipate observing relatively similar metabolites affected by similar measurement biases.
We therefore evaluate the dispersion ratio (Dratio) (Broadhurst et al. 2018) for all features in the
data set. We accomplish this task using the same function as above but
this time with the DratioFilter parameter. More filters
exist for this function and we invite the user to explore them as to
decide what is best for their dataset.
#' Compute and filter based on the Dratio
filter_dratio <- DratioFilter(threshold = 0.4,
qcIndex = res$phenotype == "QC",
studyIndex = res$phenotype != "QC",
mad = TRUE)
res <- filterFeatures(res, filter = filter_dratio, assay = "norm_imputed")The Dratio filter is a powerful tool to identify features that
exhibit high variability in the data, relating the variance observed in
QC samples with that in study samples. By setting a threshold of 0.4, we
remove features that have a high degree of variability between the QC
and study samples. In this example, any feature in which the deviation
at the QC is higher than 40% (threshold = 0.4)of the
deviation on the study samples is removed. This filtering step ensures
that only features are retained that have considerably lower technical
than biological variance.
Note that the rowDratio() and rowRsd()
functions from the MetaboCoreUtils
package could be used to calculate the actual numeric values for the
estimates used for filtering, to e.g. to evaluate their distribution in
the whole data set or identify data set-dependent threshold values.
Finally, we evaluate the number of features left after the filtering steps and calculate the percentage of features that were removed.
#' Number of features after analysis
nrow(res)## [1] 4275## [1] 47.1438The dataset has been reduced from 9068 to 4275 features. We did remove a considerable amount of features but this is expected as we want to focus on the most reliable features for our analysis. For the rest of our analysis we need to separate the QC samples from the study samples. We will store the QC samples in a separate object for later use.
res_qc <- res[, res$phenotype == "QC"]
res <- res[, res$phenotype != "QC"]We in addition calculate the CV of the QC samples and add that as an
additional column to the rowData() of our result object.
These could be used later to prioritize identified significant
features with e.g. low technical noise.
#' Calculate the QC's CV and add as feature variable to the data set
rowData(res)$qc_cv <- assay(res_qc, "norm") |>
rowRsd()Now that our data set has been preprocessed, normalized and filtered, we can start to evaluate the distribution of the data and estimate the variation due to biology.
Differential abundance analysis
After normalization and quality control, the next step is to identify features that are differentially abundant between the study groups. This crucial step allows us to identify potential biomarkers or metabolites that are associated with the study groups. There are various approaches and methods available for the identification of such features of interest. In this workflow we use a multiple linear regression analysis to identify features with significantly difference in abundances between the CVD or CTR study group.
Before performing the tests we evaluate similarities between study samples using a PCA (after excluding the QC samples to avoid them influencing the results).
col_sample <- col_phenotype[res$phenotype]
#' Log transform and scale the data for PCA analysis
vals <- assay(res, "norm_imputed") |>
t() |>
log2() |>
scale(center = TRUE, scale = TRUE)
pca_res <- prcomp(vals, scale = FALSE, center = FALSE)
vals_st <- cbind(vals, phenotype = res$phenotype)
autoplot(pca_res, data = vals_st , colour = 'phenotype', scale = 0) +
scale_color_manual(values = col_phenotype)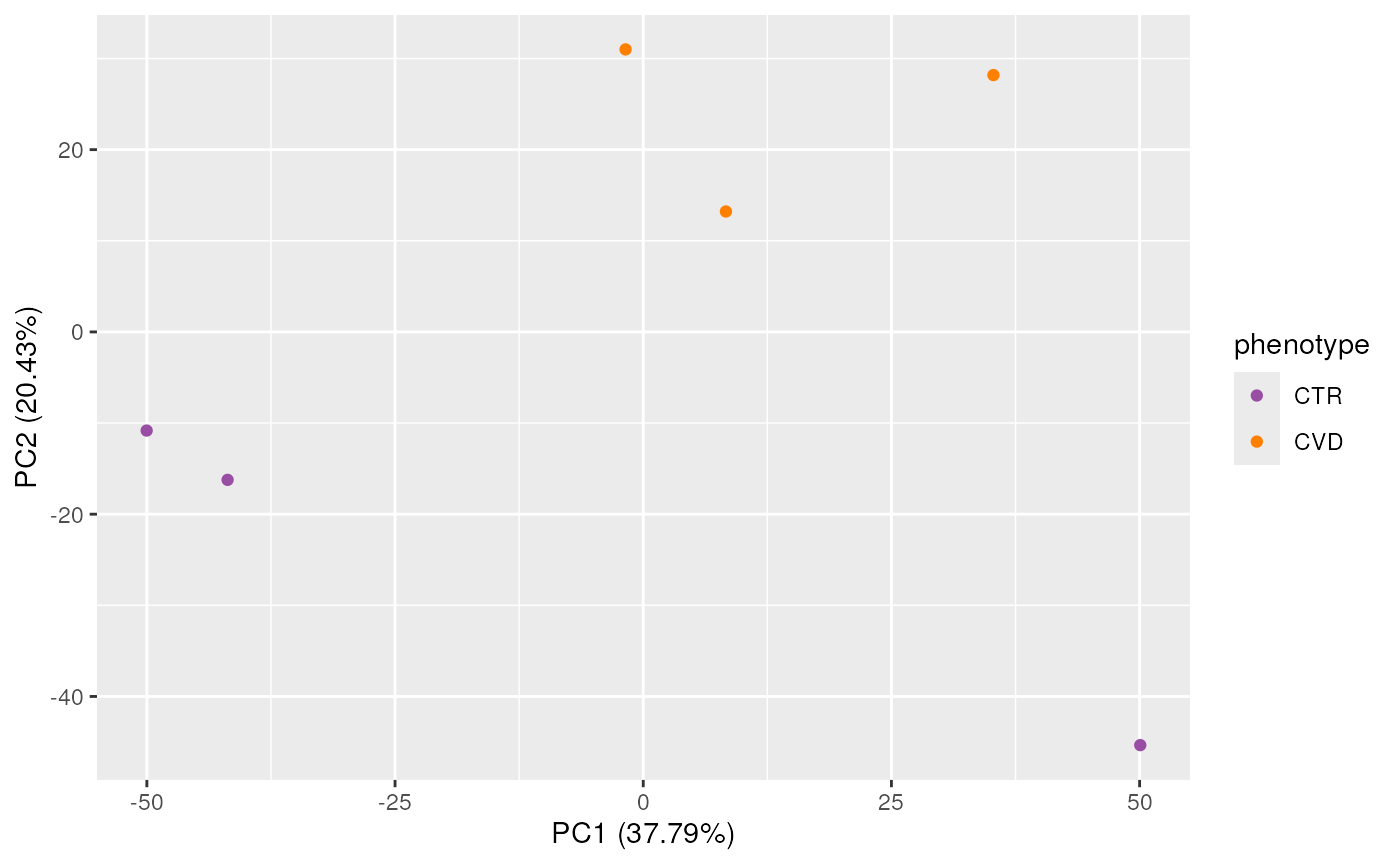
The samples clearly separate by study group on PCA indicating that there are differences in the metabolite profiles between the two groups. However, what drives the separation on PC1 is not clear. Below we evaluate whether this could be explained by the other available variable of our study, i.e., age:
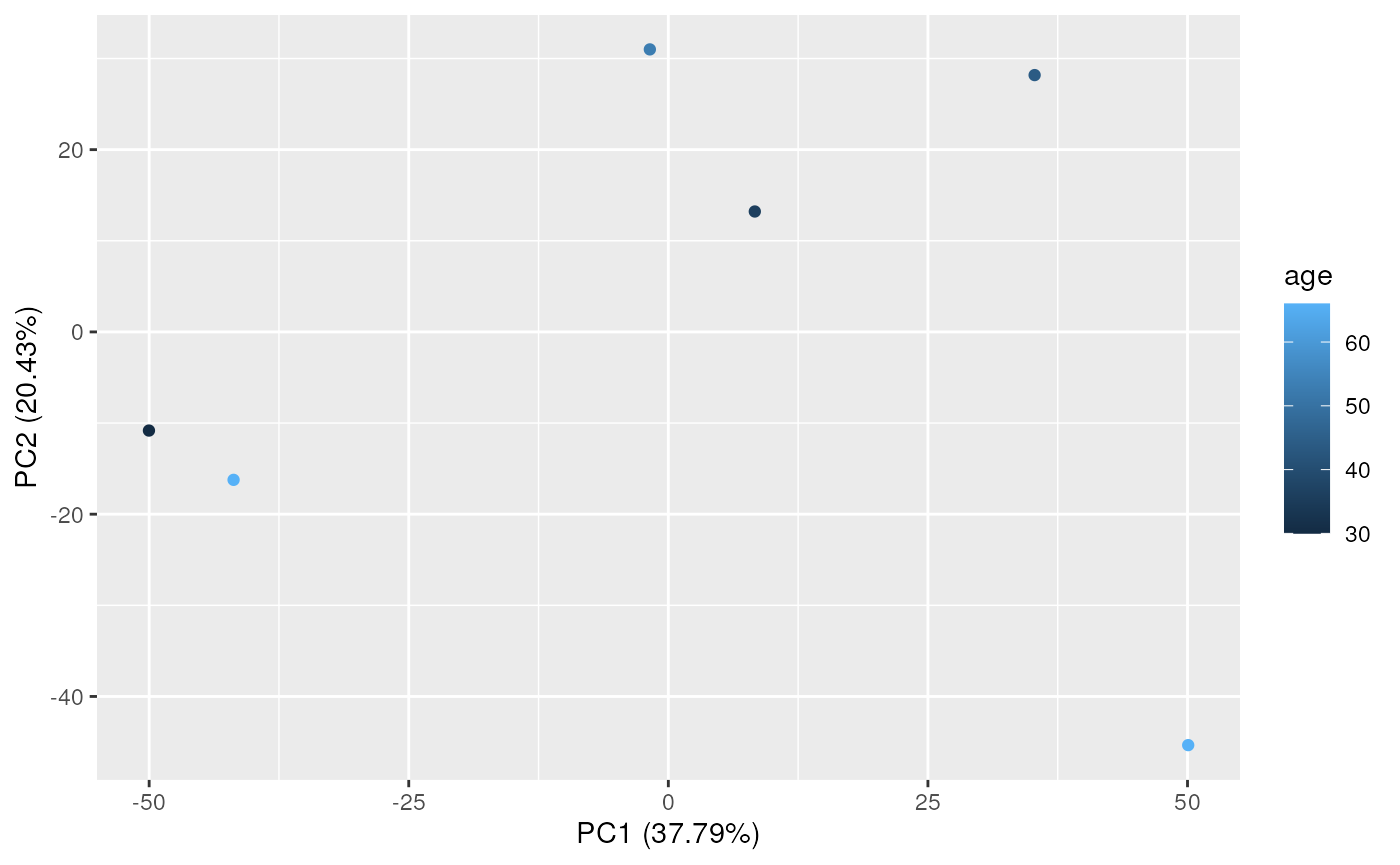
According to the PCA above, PC1 does not seem to be related to age.
Even if there is some variance in the data set we can’t explain at
this stage, we proceed with the (supervised) statistical tests to
identify features of interest. We compute linear models for each
metabolite explaining the observed feature abundance by the available
study variables. While we could also use the base R function
lm(), we utilize the R Biocpkg("limma")
package to conduct the differential abundance analysis: the
moderated test statistics (Smyth
2004) provided by this package are specifically well suited for
experiments with a limited number of replicates. For our tests we use a
linear model ~ phenotype + age, hence explaining the
abundances of one metabolite accounting for the study group assignment
and age of each individual. The analysis might benefit from inclusion of
a study covariate associated with PC2 or explaining the variance seen in
that principal component, but for the present analysis only the
participant’s age and disease association was provided.
Below we define the design of the study with the
model.matrix() function and fit feature-wise linear models
to the log2-transformed abundances using the lmFit()
function. P-values for significance of association are then calculated
using the eBayes() function, that also performs the
empirical Bayes-based robust estimation of the standard errors. See also
the excellent vignette/user guide of the limma
package for examples and details on the linear model procedure.
#' Define the linear model to be applied to the data
p.cut <- 0.05 # cut-off for significance.
m.cut <- 0.5 # cut-off for log2 fold change
age <- res$age
phenotype <- factor(res$phenotype)
design <- model.matrix(~ phenotype + age)
#' Fit the linear model to the data, explaining metabolite
#' concentrations by phenotype and age.
fit <- lmFit(log2(assay(res, "norm_imputed")), design = design)
fit <- eBayes(fit)After the linear models have been fitted, we can now proceed to
extract the results. We create a data frame containing the coefficients,
raw and adjusted p-values (applying a Benjamini-Hochberg correction,
i.e., method = "BH" for improved control of the false
discovery rate), the average intensity of signals in CVD and CTR
samples, and an indication of whether a feature is deemed significant or
not. We consider all metabolites with an adjusted p-value smaller than
0.05 to be significant, but we could also include the (absolute)
difference in abundances in the cut-off criteria. At last, we add the
differential abundance results to the result object’s
rowData().
#' Compile a result data frame
tmp <- data.frame(
coef.CVD = fit$coefficients[, "phenotypeCVD"],
pvalue.CVD = fit$p.value[, "phenotypeCVD"],
adjp.CVD = p.adjust(fit$p.value[, "phenotypeCVD"], method = "BH"),
avg.CVD = rowMeans(
log2(assay(res, "norm_imputed")[, res$phenotype == "CVD"])),
avg.CTR = rowMeans(
log2(assay(res, "norm_imputed")[, res$phenotype == "CTR"]))
)
tmp$significant.CVD <- tmp$adjp.CVD < 0.05
#' Add the results to the object's rowData
rowData(res) <- cbind(rowData(res), tmp)We can now proceed to visualize the distribution of the raw and adjusted p-values.
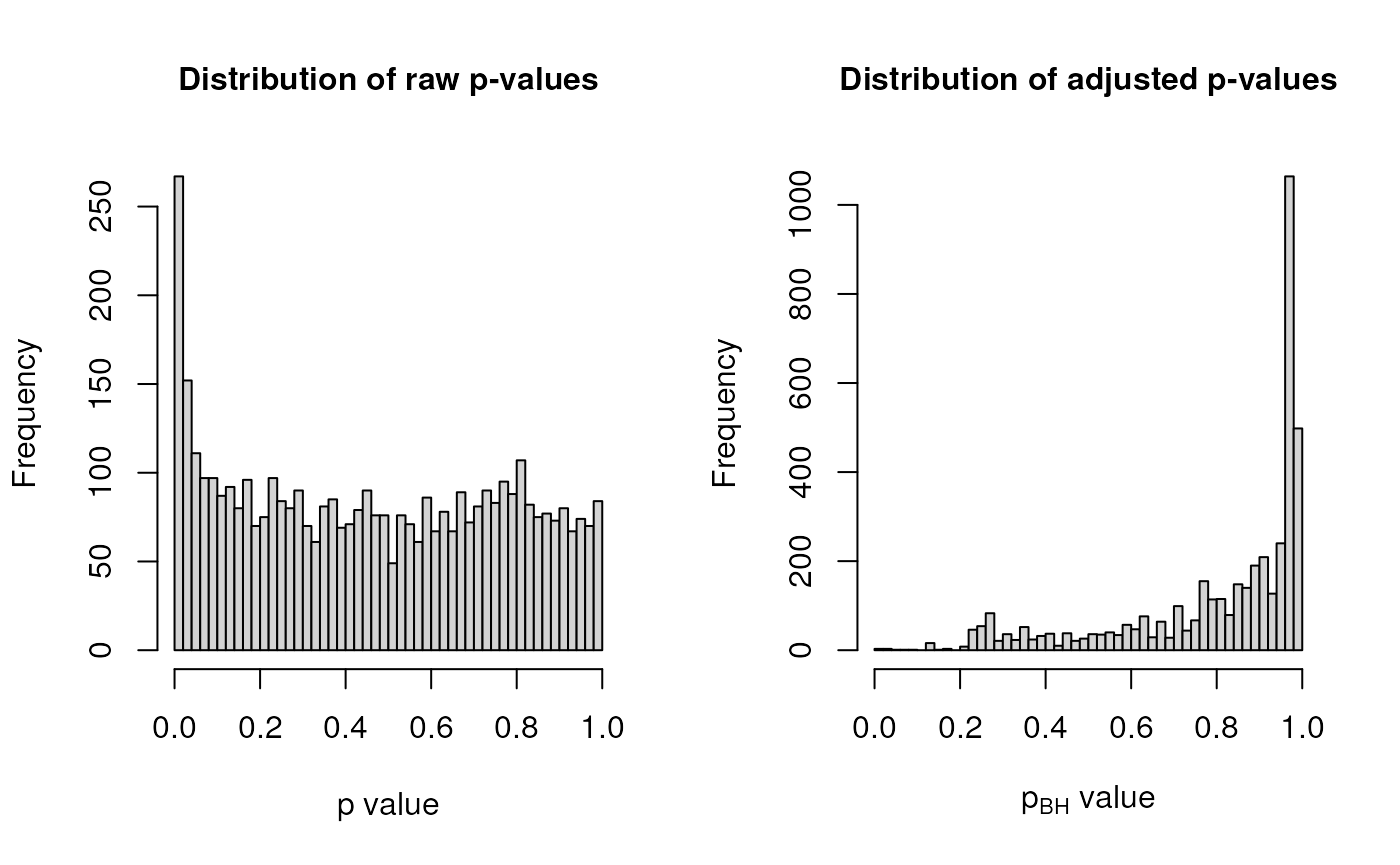
Distribution of raw (left) and adjusted p-values (right).
The histograms above show the distribution of raw and adjusted p-values. Except for an enrichment of small p-values, the raw p-values are (more or less) uniformly distributed, which indicates the absence of any strong systematic biases in the data. The adjusted p-values are more conservative and account for multiple testing; this is important here as we fit a linear model to each feature and therefore perform a large number of tests which leads to a high chance of false positive findings. We do see that some features have very low p-values, indicating that they are likely to be significantly different between the two study groups.
Below we plot the adjusted p-values against the log2 fold change of (average) abundances. This volcano plot will allow us to visualize the features that are significantly different between the two study groups. These are highlighted with a blue color in the plot below.
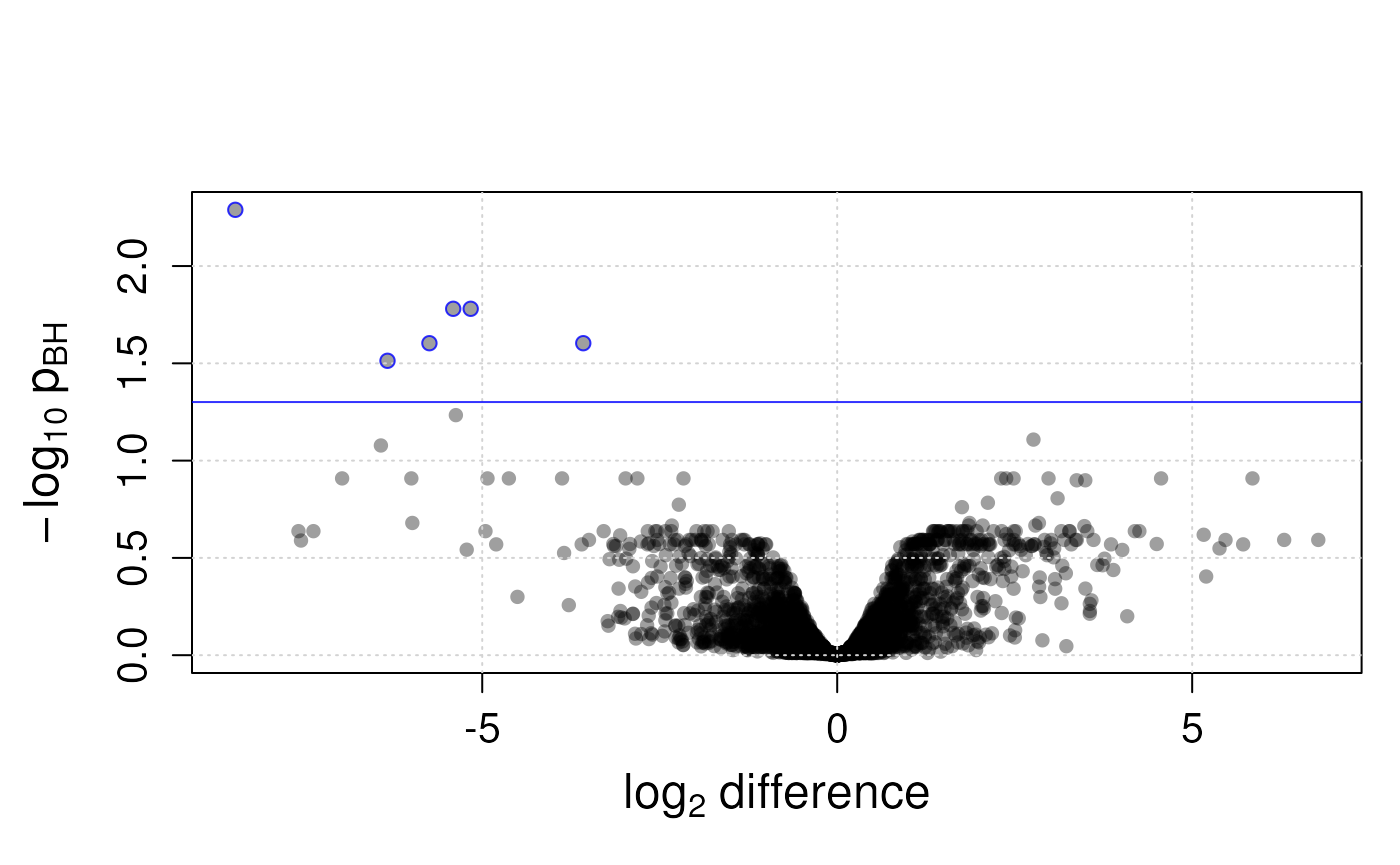
Volcano plot showing the analysis results.
The most interesting features are in the top corners in the volcano plot (i.e., features with a large difference in abundance between the groups and a small p-value). All significant features have a negative coefficient (log2 fold change value) indicating that their abundance is lower in CVD samples compared to the CTR samples. These features are listed, along with their average difference in (log2) abundance between the compared groups, their adjusted p-values, their average (log2) abundance in each sample group and their RSD (CV) in QC samples in the table below.
| mzmed | rtmed | coef.CVD | adjp.CVD | avg.CTR | avg.CVD | |
|---|---|---|---|---|---|---|
| FT0732 | 182.1 | 34.84 | -8.478 | 0.00514 | 12.23 | 3.866 |
| FT0845 | 195.1 | 32.66 | -6.334 | 0.03068 | 16.9 | 10.45 |
| FT0565 | 161 | 162.1 | -5.744 | 0.02492 | 10.28 | 4.297 |
| FT1171 | 229.1 | 181.1 | -5.409 | 0.01658 | 10.72 | 5.5 |
| FT0371 | 138.1 | 148.4 | -5.163 | 0.01658 | 9.912 | 4.448 |
| FT5606 | 560.4 | 33.55 | -3.577 | 0.02492 | 8.883 | 5.111 |
| qc_cv | |
|---|---|
| FT0732 | 0.2103 |
| FT0845 | 0.02961 |
| FT0565 | 0.03597 |
| FT1171 | 0.07075 |
| FT0371 | 0.5556 |
| FT5606 | 1.215 |
Below we visualize the EICs of the significant features to evaluate their (raw) signal. We restrict the MS data set to study samples. Parameters
-
keepFeatures = TRUE: ensures that identified features are retained in the `subset object. -
peakBg: defines the (background) color for each individual chromatographic peak in the EIC object.
#' Restrict the raw data to study samples.
data_study <- data[sampleData(data)$phenotype != "QC", keepFeatures = TRUE]
#' Extract EICs for the significant features
eic_sign <- featureChromatograms(
data_study, features = rownames(tab), expandRt = 5, filled = TRUE)
#' Plot the EICs.
plot(eic_sign, col = col_sample,
peakBg = paste0(col_sample[chromPeaks(eic_sign)[, "sample"]], 40))
legend("topright", col = col_phenotype, legend = names(col_phenotype), lty = 1)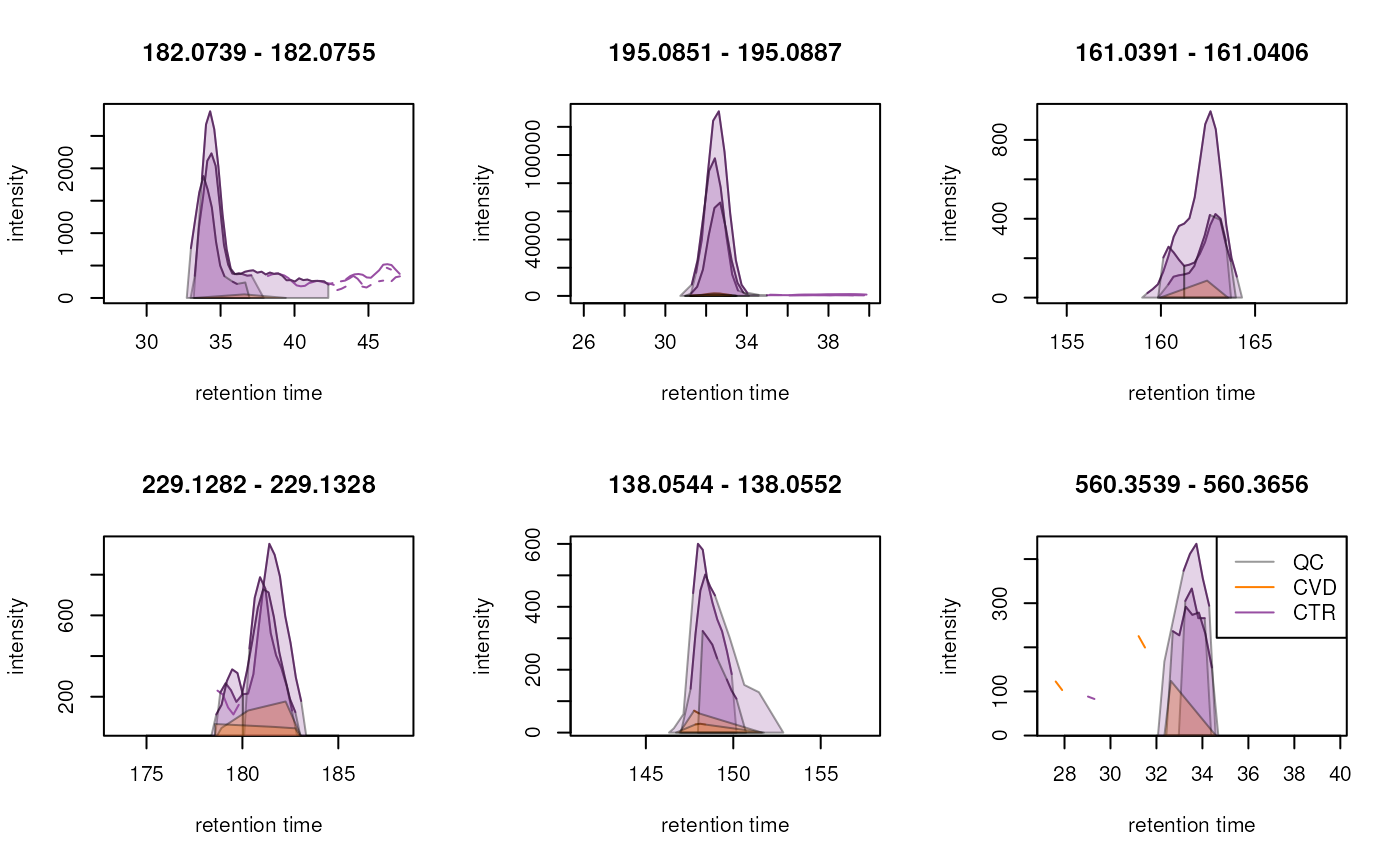
The EICs for all significant features show a clear and single peak. Their intensities are (as already observed above) much larger in the CTR than in the CVD samples. With the exception of the second feature (second EIC in the top row), the intensities for all significant features are however generally low. This might make it challenging to identify them using an LC-MS/MS setup.
Annotation
We have now identified the features with significant differences in abundances between the two study groups. These provide information on metabolic pathways that differentiate affected from healthy individuals and might hence also serve as biomarkers. However, at this stage of the analysis we do not know what compounds/metabolites they actually represent. We thus need now to annotate these signals. Annotation can be performed at different level of confidence Schymanski et al. (2014). The lowest level of annotation, with the highest rate of false positive hits, bases on the features m/z ratios. Higher levels of annotations employ fragment spectra (MS2 spectra) of ions of interest requiring this however acquisition of additional data. In this section, we will demonstrate multiple ways to annotate our significant features using functionality provided through Bioconductor packages. Alternative approaches and external software tools, which may be better suited, will also be discussed later in the section.
MS1-based annotation
Our data set was acquired using an LC-MS setup and our features are thus only characterized by their m/z and retention times. The retention time is LC-setup-specific and, without prior data/knowledge provide only little information on the features’ identity. Modern MS instruments have a high accuracy and the m/z values should therefore be reliable estimates of the compound ion’s mass-to-charge ratio. As first approach, we use the features’ m/z values and match them against reference values, i.e., exact masses of chemical compounds that are provided by a reference database, in our case the MassBank database. The full MassBank data is re-distributed through Bioconductor’s AnnotationHub resource, which simplifies their integration into reproducible R-based analysis workflows.
Below we load the resource, list all available MassBank data sets/releases and load one of them.
#' load reference data
ah <- AnnotationHub()
#' List available MassBank data sets
query(ah, "MassBank")## AnnotationHub with 6 records
## # snapshotDate(): 2024-08-01
## # $dataprovider: MassBank
## # $species: NA
## # $rdataclass: CompDb
## # additional mcols(): taxonomyid, genome, description,
## # coordinate_1_based, maintainer, rdatadateadded, preparerclass, tags,
## # rdatapath, sourceurl, sourcetype
## # retrieve records with, e.g., 'object[["AH107048"]]'
##
## title
## AH107048 | MassBank CompDb for release 2021.03
## AH107049 | MassBank CompDb for release 2022.06
## AH111334 | MassBank CompDb for release 2022.12.1
## AH116164 | MassBank CompDb for release 2023.06
## AH116165 | MassBank CompDb for release 2023.09
## AH116166 | MassBank CompDb for release 2023.11
#' Load one MAssBank release
mb <- ah[["AH116166"]]The MassBank data is provided as a self-contained SQLite database and
data can be queried and accessed through the CompoundDb
Bioconductor package. Below we use the compounds() function
to extract small compound annotations from the database.
cmps <- compounds(mb, columns = c("compound_id", "name", "formula",
"exactmass", "inchikey"))
head(cmps)## compound_id formula exactmass inchikey
## 1 1 C27H29NO11 543.1741 AOJJSUZBOXZQNB-UHFFFAOYSA-N
## 2 2 C40H54O4 598.4022 KFNGKYUGHHQDEE-AXWOCEAUSA-N
## 3 3 C10H24N2O2 204.1838 AEUTYOVWOVBAKS-UWVGGRQHSA-N
## 4 4 C16H27NO5 313.1889 LMFKRLGHEKVMNT-UJDVCPFMSA-N
## 5 5 C20H15Cl3N2OS 435.9971 JLGKQTAYUIMGRK-UHFFFAOYSA-N
## 6 6 C15H14O5 274.0841 BWNCKEBBYADFPQ-UHFFFAOYSA-N
## name
## 1 Epirubicin
## 2 Crassostreaxanthin A
## 3 Ethambutol
## 4 Heliotrine
## 5 Sertaconazole
## 6 (R)SemivioxanthinMassBank (as most other small compound annotation databases) provides
the (exact) molecular mass of each compound. Since almost all small
compounds are neutral in their natural state, they need to be first
converted to m/z values to allow matching against the feature’s
m/z. To calculate an m/z from a neutral mass, we need
to assume which ion (adduct) might be generated from the measured
metabolites by the employed electro-spray ionization. In positive
polarity, for human serum samples, the most common ions will be
protonated ([M+H]+), or bear the addition of sodium
([M+Na]+) or ammonium ([M+H-NH3]+) ions. To match the
observed m/z values against reference values from these
potential ions we use again the matchValues() function with
the Mass2MzParam approach, that allows to specify the types
of expected ions with its adducts parameter and the maximal
allowed difference between the compared values using the
tolerance and ppm parameters. Below we first
prepare the data.frame with the significant features, set up the
parameters for the matching and perform the comparison of the query
features against the reference database.
#' Prepare query data frame
rowData(res)$feature_id <- rownames(rowData(res))
res_sig <- res[rowData(res)$significant.CVD, ]
#' Setup parameters for the matching
param <- Mass2MzParam(adducts = c("[M+H]+", "[M+Na]+", "[M+H-NH3]+"),
tolerance = 0, ppm = 5)
#' Perform the matching.
mtch <- matchValues(res_sig, cmps, param = param, mzColname = "mzmed")
mtch## Object of class Matched
## Total number of matches: 237
## Number of query objects: 6 (4 matched)
## Number of target objects: 117732 (237 matched)The resulting Matched object shows that 4 of our 6
significant features could be matched to ions of compounds from the
MassBank database. Below we extract the full result from the
Matched object.
#' Extracting the results
mtch_res <- matchedData(mtch, c("feature_id", "mzmed", "rtmed",
"adduct", "ppm_error",
"target_formula", "target_name",
"target_inchikey"))
mtch_res## DataFrame with 239 rows and 8 columns
## feature_id mzmed rtmed adduct ppm_error target_formula
## <character> <numeric> <numeric> <character> <numeric> <character>
## FT0371 FT0371 138.055 148.396 [M+H]+ 2.08055 C7H7NO2
## FT0371 FT0371 138.055 148.396 [M+H]+ 1.93568 C7H7NO2
## FT0371 FT0371 138.055 148.396 [M+H]+ 1.93568 C7H7NO2
## FT0371 FT0371 138.055 148.396 [M+H]+ 1.93568 C7H7NO2
## FT0371 FT0371 138.055 148.396 [M+H]+ 2.08055 C7H7NO2
## ... ... ... ... ... ... ...
## FT1171 FT1171 229.13 181.0883 [M+Na]+ 3.07708 C12H18N2O
## FT1171 FT1171 229.13 181.0883 [M+Na]+ 3.07708 C12H18N2O
## FT1171 FT1171 229.13 181.0883 [M+Na]+ 3.07708 C12H18N2O
## FT1171 FT1171 229.13 181.0883 [M+Na]+ 3.07708 C12H18N2O
## FT5606 FT5606 560.36 33.5492 NA NA NA
## target_name target_inchikey
## <character> <character>
## FT0371 Benzohydro... VDEUYMSGMP...
## FT0371 Trigonelli... WWNNZCOKKK...
## FT0371 Trigonelli... WWNNZCOKKK...
## FT0371 Trigonelli... WWNNZCOKKK...
## FT0371 Salicylami... SKZKKFZAGN...
## ... ... ...
## FT1171 Isoproturo... PUIYMUZLKQ...
## FT1171 Isoproturo... PUIYMUZLKQ...
## FT1171 Isoproturo... PUIYMUZLKQ...
## FT1171 Isoproturo... PUIYMUZLKQ...
## FT5606 NA NAThus, in total 237 ions of compounds in MassBank were matched to our significant features based on the specified tolerance settings. Many compounds, while having a different structure and thus function/chemical property, have an identical chemical formula and thus mass. Matching exclusively on the m/z of features will hence result in many potentially false positive hits and is thus considered to provide only low confidence annotation. An additional complication is that some annotation resources, like MassBank, being community maintained, contain a large amount of redundant information. To reduce redundancy in the result table we below iterate over the hits of a feature and keep only matches to unique compounds (identified by their INCHIKEY). The INCHI or INCHIKEY combine information from compound’s chemical formula and structure, so while different compounds can share the same chemical formula, they should have a different structure and thus INCHI.
rownames(mtch_res) <- NULL
#' Keep only info on features that machted - create a utility function for that
mtch_res <- split(mtch_res, mtch_res$feature_id) |>
lapply(function(x) {
lapply(split(x, x$target_inchikey), function(z) {
z[which.min(z$ppm_error), ]
})
}) |>
unlist(recursive = FALSE) |>
do.call(what = rbind)
#' Display the results
mtch_res |>
as.data.frame() |>
pandoc.table(style = "rmarkdown", caption = "Table 8.MS1 annotation results")| feature_id | mzmed | rtmed | adduct | ppm_error | target_formula |
|---|---|---|---|---|---|
| FT0371 | 138.1 | 148.4 | [M+H]+ | 1.936 | C7H7NO2 |
| FT0371 | 138.1 | 148.4 | [M+H]+ | 1.936 | C7H7NO2 |
| FT0371 | 138.1 | 148.4 | [M+H]+ | 1.936 | C7H7NO2 |
| FT0371 | 138.1 | 148.4 | [M+H]+ | 1.936 | C7H7NO2 |
| FT0371 | 138.1 | 148.4 | [M+H]+ | 1.936 | C7H7NO2 |
| FT0371 | 138.1 | 148.4 | [M+H]+ | 1.936 | C7H7NO2 |
| FT0371 | 138.1 | 148.4 | [M+H]+ | 1.925 | C7H7NO2 |
| FT0371 | 138.1 | 148.4 | [M+H]+ | 1.936 | C7H7NO2 |
| FT0371 | 138.1 | 148.4 | [M+H]+ | 1.936 | C7H7NO2 |
| FT0371 | 138.1 | 148.4 | [M+H]+ | 1.936 | C7H7NO2 |
| FT0371 | 138.1 | 148.4 | [M+H]+ | 1.925 | C7H7NO2 |
| FT0371 | 138.1 | 148.4 | [M+H]+ | 1.936 | C7H7NO2 |
| FT0371 | 138.1 | 148.4 | [M+H]+ | 1.936 | C7H7NO2 |
| FT0371 | 138.1 | 148.4 | [M+H]+ | 1.936 | C7H7NO2 |
| FT0371 | 138.1 | 148.4 | [M+H]+ | 1.936 | C7H7NO2 |
| FT0371 | 138.1 | 148.4 | [M+H]+ | 1.936 | C7H7NO2 |
| FT0565 | 161 | 162.1 | [M+Na]+ | 3.125 | C8H10S |
| FT0845 | 195.1 | 32.66 | [M+H]+ | 0.06147 | C8H10N4O2 |
| FT0845 | 195.1 | 32.66 | [M+H]+ | 0.06147 | C8H10N4O2 |
| FT0845 | 195.1 | 32.66 | [M+H]+ | 0.1866 | C8H10N4O2 |
| FT1171 | 229.1 | 181.1 | [M+Na]+ | 3.077 | C12H18N2O |
| target_name | target_inchikey |
|---|---|
| 4-Aminobenzoic acid | ALYNCZNDIQEVRV-UHFFFAOYSA-N |
| 2-PYRIDINECARBOXYLIC ACID METHYL ESTER | NMMIHXMBOZYNET-UHFFFAOYSA-N |
| 4-PYRIDINECARBOXYLIC ACID METHYL ESTER | OLXYLDUSSBULGU-UHFFFAOYSA-N |
| SALICYLALDOXIME | ORIHZIZPTZTNCU-VMPITWQZSA-N |
| ORTHO NITROTOLUENE | PLAZTCDQAHEYBI-UHFFFAOYSA-N |
| 4-PYRIDYL ACETATE | PTZQTYADHHANGW-UHFFFAOYSA-N |
| Salicylaldehyde oxime | PUNVNOQWQQPNES-UHFFFAOYSA-N |
| META NITROTOLUENE | QZYHIOPPLUPUJF-UHFFFAOYSA-N |
| Anthranilic acid | RWZYAGGXGHYGMB-UHFFFAOYSA-N |
| 2-HYDROXYBENZAMIDE | SKZKKFZAGNVIMN-UHFFFAOYSA-N |
| N-Hydroxybenzamide | VDEUYMSGMPQMIK-UHFFFAOYSA-N |
| 3-Pyridylacetic acid | WGNUNYPERJMVRM-UHFFFAOYSA-N |
| Trigonelline | WWNNZCOKKKDOPX-UHFFFAOYSA-N |
| META-AMINOBENZOIC ACID | XFDUHJPVQKIXHO-UHFFFAOYSA-N |
| 3-PYRIDINECARBOXYLIC ACID METHYL ESTER | YNBADRVTZLEFNH-UHFFFAOYSA-N |
| 4-NITROTOLUENE | ZPTVNYMJQHSSEA-UHFFFAOYSA-N |
| BENZYL METHYL SULFIDE | OFQPKKGMNWASPN-UHFFFAOYSA-N |
| Isocaffeine | LPHGQDQBBGAPDZ-UHFFFAOYSA-N |
| Caffeine | RYYVLZVUVIJVGH-UHFFFAOYSA-N |
| 1,3-Benzenedicarboxylic acid, dihydrazide | UTTHLMXOSUFZCQ-UHFFFAOYSA-N |
| Isoproturon | PUIYMUZLKQOUOZ-UHFFFAOYSA-N |
The table above shows the results of the MS1-based annotation process. We can see that four of our significant features were matched. The matches seem to be pretty accurate with low ppm errors. The deduplication performed above considerably reduced the number of hits for each feature, but the first still matches ions of a large number of compounds (all with the same chemical formula).
Considering both features’ m/z and retention times in the MS1-based annotation will increase the annotation confidence, but requires additional data, such as recording of the retention time of thepure standard for a compound on the same LC setup. An alternative approach which might provide better inside on annotations and help to choose between different annotations for a same feature is to evaluate certain chemical properties of the possible matches. For instance, the LogP value, available in several databases such HMDB, provides an insight on a given compound’s polarity. As this property highly affects the interaction of the analyte with the column, it usually also directly affects separation. Therefore, a comparison between an analyte’s retention time and polarity can help to rule out some possible misidentifications.
While being of low confidence, MS1-based annotation can provide first candidate annotations that could be confirmed or rejected by additional analyses.
MS2-based annotation
MS1 annotation is a fast and efficient method to annotate features and therefore give a first insight into the compounds that are significantly different between the two study groups. However, it is not always the most accurate. MS2 data can provide a higher level of confidence in the annotation process as it provides, through the observed fragmentation pattern, information on the structure of the compound.
MS2 data can be generated through LC-MS/MS measurement in which MS2 spectra are recorded for ions either in a data dependent acquisition (DDA) or data independent acquisition (DIA) mode. Generally, it is advised to include some LC-MS/MS runs of QC samples or randomly selected study samples already during the acquisition of the MS1 data that is used for quantification of the signals. As an alternative, or in addition, a post-hoc LC-MS/MS acquisition can be performed to generate the MS2 data needed for annotation. For the present experiment, a separate LC-MS/MS measurement was conducted on QC samples and selected study samples to generate such data using an inclusion list of pre-selected ions. These represent features found to be significantly different between CVD and CTR samples in an initial analysis of the full experiment. We use a subset of this second LC-MS/MS data set to show how such data can be used for MS2-based annotation. In our differential abundance analysis we found features with significantly higher abundances in CTR samples. Consequently, we will utilize the MS2 data obtained from the CTR samples to annotate these significant features.
Below we load the LC-MS/MS data from the experiment and restrict it to data acquired from the CTR sample.
#' Load form the MetaboLights Database
param <- MetaboLightsParam(mtblsId = "MTBLS8735",
assayName = paste0("a_MTBLS8735_LC-MSMS_positive_",
"hilic_metabolite_profiling.txt"),
filePattern = ".mzML")
msms_data <- readMsObject(MsExperiment(),
param,
keepOntology = FALSE,
keepProtocol = FALSE,
simplify = TRUE)
#adjust sampleData
colnames(sampleData(msms_data)) <- c("sample_name", "derived_spectra_data_file",
"metabolite_asssignment_file",
"source_name",
"organism",
"blood_sample_type",
"sample_type", "age", "unit", "phenotype")
# filter samples to keep MSMS data from CTR samples:
sampleData(msms_data) <- sampleData(msms_data)[sampleData(msms_data)$phenotype == "CTR", ]
sampleData(msms_data) <- sampleData(msms_data)[grepl("MSMS", sampleData(msms_data)$derived_spectra_data_file), ]
# Add fragmentation data information (from filenames)
sampleData(msms_data)$fragmentation_mode <- c("CE20", "CE30", "CES")
#let's look at the updated sample data
sampleData(msms_data)[, c("derived_spectra_data_file",
"phenotype", "sample_name", "age")] |>
as.data.frame() |>
pandoc.table(style = "rmarkdown",
caption = "Table 1. Samples from the data set.")##
##
## | derived_spectra_data_file | phenotype | sample_name | age |
## |:----------------------------:|:---------:|:-----------:|:---:|
## | FILES/MSMS_2_E_CE20_POS.mzML | CTR | E | 66 |
## | FILES/MSMS_2_E_CE30_POS.mzML | CTR | E | 66 |
## | FILES/MSMS_2_E_CES_POS.mzML | CTR | E | 66 |
##
## Table: Table 1. Samples from the data set.
#' Filter the data to the same RT range as the LC-MS run
msms_data <- filterRt(msms_data, c(10, 240))In total we have 3 LC-MS/MS data files for the control samples, each with a different collision energy to fragment the ions. Below we show the number of MS1 and MS2 spectra for each of the files.
#' check the number of spectra per ms level
spectra(msms_data) |>
msLevel() |>
split(spectraSampleIndex(msms_data)) |>
lapply(table) |>
do.call(what = cbind)## 1 2 3 4 5 6 7 8 9 10 11 12
## 1 825 186 186 186 825 186 186 186 825 185 186 185
## 2 825 3121 3118 3124 825 3123 3118 3120 825 3117 3117 3116Compared to the number of MS2 spectra, far less MS1 spectra were acquired. The configuration of the MS instrument was set to ensure that the ions specified in the inclusion list were selected for fragmentation, even if their intensity might be very low. With this setting, however, most of the recorded MS2 spectra will represent only noise. The plot below shows the location of precursor ions in the m/z - retention time plane for the three files.
plotPrecursorIons(msms_data)
We can see MS2 spectra being recorded for the m/z of interest along the full retention time range, even if the actual ions will only be eluting within certain retention time windows.
We next extract the Spectra object with the MS data from
the data object and assign a new spectra variable with the employed
collision energy, which we extract from the data object
sampleData.
ms2_ctr <- spectra(msms_data)
ms2_ctr$collision_energy <-
sampleData(msms_data)$fragmentation_mode[spectraSampleIndex(msms_data)]We next filter the MS data by first restricting to MS2 spectra and then removing mass peaks from each spectrum with an intensity that is lower than 5% of the highest intensity for that spectrum, assuming that these very low intensity peaks represent background signal.
#' Remove low intensity peaks
low_int <- function(x, ...) {
x > max(x, na.rm = TRUE) * 0.05
}
ms2_ctr <- filterMsLevel(ms2_ctr, 2L) |>
filterIntensity(intensity = low_int)We next remove also mass peaks with an m/z value greater or equal to the precursor m/z of the ion. This puts, for the later matching against reference spectra, more weight on the fragmentation pattern of ions and avoids hits based only on the precursor m/z peak (and hence a similar mass of the compared compounds). At last, we restrict the data to spectra with at least two fragment peaks and scale their intensities such that their sum is 1 for each spectrum. While similarity calculations are not affected by this scaling, it makes visual comparison of fragment spectra easier to read.
#' Remove precursor peaks and restrict to spectra with a minimum
#' number of peaks
ms2_ctr <- filterPrecursorPeaks(ms2_ctr, ppm = 50, mz = ">=")
ms2_ctr <- ms2_ctr[lengths(ms2_ctr) > 1] |>
scalePeaks()Finally, also to speed up the later comparison of the spectra against
the reference database, we load the full MS data into memory (by
changing the backend to MsBackendMemory) and
apply all processing steps performed on this data so far.
Keeping the MS data in memory has performance benefits, but is generally
not suggested for large data sets. To evaluate the impact for the
present data set we print in addition the size of the data object before
and after changing the backend.
#' Size of the object before loading into memory
print(object.size(ms2_ctr), units = "MB")## 5.1 Mb
#' Load the MS data subset into memory
ms2_ctr <- setBackend(ms2_ctr, MsBackendMemory())
ms2_ctr <- applyProcessing(ms2_ctr)
#' Size of the object after loading into memory
print(object.size(ms2_ctr), units = "MB")## 18.2 MbThere is thus only a moderate increase in memory demand after loading the MS data into memory (also because we filtered and cleaned the MS2 data).
While we could proceed and match all these experimental MS2 spectra
against reference fragment spectra, for our workflow we aim to annotate
the features found significant in the differential abundance analysis.
The goal is thus to identify the MS2 spectra from the second (LC-MS/MS)
run that could represent fragments of the ions of features from the data
in the first (LC-MS) run. Our approach is to match MS2 spectra against
the significant features determined earlier based on their precursor
m/z and retention time (given an acceptable tolerance) to the
feature’s m/z and retention time. This can be easily done using
the featureArea() function that effectively considers the
actual m/z and retention time ranges of the features’
chromatographic peaks and therefore increase the chance of finding a
correct match. This however also assumes that retention times between
the first and second run don’t differ by much. Alternatively, we would
need to align the retention times of the second LC-MS/MS data set to
those of the first.
Below we first extract the feature area, i.e., the m/z and retention time ranges, for the significant features.
#' Define the m/z and retention time ranges for the significant features
target <- featureArea(data)[rownames(res_sig), ]
target## mzmin mzmax rtmin rtmax
## FT0371 138.0544 138.0552 146.32270 152.86115
## FT0565 161.0391 161.0407 159.00234 164.30799
## FT0732 182.0726 182.0756 32.71242 42.28755
## FT0845 195.0799 195.0887 30.73235 35.67337
## FT1171 229.1282 229.1335 178.01450 183.35303
## FT5606 560.3539 560.3656 32.06570 35.33456We next identify the fragment spectra with their precursor
m/z and retention times within these ranges. We use the
filterRanges() function that allows to filter a
Spectra object using multiple ranges simultaneously. We
apply this function separately to each feature (row in the above matrix)
to extract the MS2 spectra representing fragmentation information of the
presumed feature’s ions.
#' Identify for each feature MS2 spectra with their precursor m/z and
#' retention time within the feature's m/z and retention time range
ms2_ctr_fts <- apply(target[, c("rtmin", "rtmax", "mzmin", "mzmax")],
MARGIN = 1, FUN = filterRanges, object = ms2_ctr,
spectraVariables = c("rtime", "precursorMz"))
lengths(ms2_ctr_fts)## FT0371 FT0565 FT0732 FT0845 FT1171 FT5606
## 38 36 135 68 38 0The result from this apply() call is a list
of Spectra, each element representing the result for one
feature. With the exception of the last feature, multiple MS2 spectra
could be identified. We next combine the list of
Spectra into a single Spectra object using the
concatenateSpectra() function and add an additional spectra
variable containing the respective feature identifier.
l <- lengths(ms2_ctr_fts)
#' Combine the individual Spectra objects
ms2_ctr_fts <- concatenateSpectra(ms2_ctr_fts)
#' Assign the feature identifier to each MS2 spectrum
ms2_ctr_fts$feature_id <- rep(rownames(res_sig), l)We have now a Spectra object with fragment spectra for
the significant features from our differential expression analysis.
We next build our reference data which we need to process the same way as our query spectra. We extract all fragment spectra from the MassBank database, restrict to positive polarity data (since our experiment was acquired in positive polarity) and perform the same processing to the fragment spectra from the MassBank database.
ms2_ref <- Spectra(mb) |>
filterPolarity(1L) |>
filterIntensity(intensity = low_int) |>
filterPrecursorPeaks(ppm = 50, mz = ">=")
ms2_ref <- ms2_ref[lengths(ms2_ref) > 1] |>
scalePeaks()Note that here we could switch to a MsBackendMemory
backend hence loading the full data from the reference database into
memory. This could have a positive impact on the performance of the
subsequent spectra matching, however it would also increase the memory
demand of the present analysis.
Now that both the Spectra object for the second run and
the database spectra have been prepared, we can proceed with the
matching process. We use the matchSpectra() function from
the MetaboAnnotation
package with the CompareSpectraParam to define the settings
for the matching. With the following parameters:
-
requirePrecursor = TRUE: Limits spectra similarity calculations to fragment spectra with a similar precursor m/z. -
toleranceandppm: Defines the acceptable difference between compared m/z values. These relaxed tolerance settings ensure that we find matches even if reference spectra were acquired on instruments with a lower accuracy. -
THRESHFUN: Defines which matches to report. Here, we keep all matches resulting in a spectra similarity score (calculated with the normalized dot product (Stein and Scott 1994), the default similarity function) larger than 0.6.
register(SerialParam())
#' Define the settings for the spectra matching.
prm <- CompareSpectraParam(ppm = 40, tolerance = 0.05,
requirePrecursor = TRUE,
THRESHFUN = function(x) which(x >= 0.6))
ms2_mtch <- matchSpectra(ms2_ctr_fts, ms2_ref, param = prm)
ms2_mtch## Object of class MatchedSpectra
## Total number of matches: 214
## Number of query objects: 315 (16 matched)
## Number of target objects: 69561 (21 matched)Thus, from the total 315 query MS2 spectra, only 16 could be matched to (at least) one reference fragment spectrum.
Below we restrict the results to these matching spectra and extract
all metadata of the query and target spectra as well as the similarity
score (the complete list of available metadata information can be listed
with the colnames() function).
#' Keep only query spectra with matching reference spectra
ms2_mtch <- ms2_mtch[whichQuery(ms2_mtch)]
#' Extract the results
ms2_mtch_res <- matchedData(ms2_mtch)
nrow(ms2_mtch_res)## [1] 214Now, we have query-target pairs with a spectra similarity higher than 0.6. Similar to the MS1-based annotation also this result table contains redundant information: we have multiple fragment spectra per feature and also MassBank contains several fragment spectra for each compound, measured using differing collision energies or MS instruments, by different laboratories. Below we thus iterate over the feature-compound pairs and select for each the one with the highest score. As an identifier for a compound, we use its fragment spectra’s INCHI-key, since compound names in MassBank do not have accepted consensus/controlled vocabularies.
#' - split the result per feature
#' - select for each feature the best matching result for each compound
#' - combine the result again into a data frame
ms2_mtch_res <-
ms2_mtch_res |>
split(f = paste(ms2_mtch_res$feature_id, ms2_mtch_res$target_inchikey)) |>
lapply(function(z) {
z[which.max(z$score), ]
}) |>
do.call(what = rbind) |>
as.data.frame()
#' List the best matching feature-compound pair
pandoc.table(ms2_mtch_res[, c("feature_id", "target_name", "score",
"target_inchikey")],
style = "rmarkdown",
caption = "Table 9.MS2 annotation results.",
split.table = Inf)| feature_id | target_name | score | target_inchikey |
|---|---|---|---|
| FT0371 | 3-Dimethylaminophenol | 0.6274 | MESJRHHDBDCQTH-UHFFFAOYSA-N |
| FT0371 | 2-Methoxy-5-methylaniline | 0.6149 | WXWCDTXEKCVRRO-UHFFFAOYSA-N |
| FT0845 | Isocaffeine | 0.8048 | LPHGQDQBBGAPDZ-UHFFFAOYSA-N |
| FT0845 | Caffeine | 0.9205 | NA |
| FT0845 | Caffeine | 0.9998 | RYYVLZVUVIJVGH-UHFFFAOYSA-N |
Thus, from the 6 significant features, only one could be annotated to a compound based on the MS2-based approach. There could be many reasons for the failure to find matches for the other features. Although MS2 spectra were selected for each feature, most appear to only represent noise, as for most features, in the LC-MS/MS run, no or only very low MS1 signal was recorded, indicating that in that selected sample the original compound might not (or no longer) be present. Also, most reference databases contain predominantly fragment spectra for protonated ([M+H]+) ions of compounds, while some of the features might represent signal from other types of ions which would result in a different fragmentation pattern. Finally, fragment spectra of compounds of interest might also simply not be present in the used reference database.
Thus, combining the information from the MS1- and MS2 based annotation we can only annotate one feature with a considerable confidence. The feature with the m/z of 195.0879 and a retention time of 32 seconds seems to be an ion of caffeine. While the result is somewhat disappointing it also clearly shows the importance of proper experimental planning and the need to control for all potential confounding factors. For the present experiment, no disease-specific biomarker could be identified, but a life-style property of individuals suffering from this disease: coffee consumption was probably contraindicated to patients of the CVD group to reduce the risk of heart arrhythmia.
We below plot the EIC for this feature highlighting the retention time at which the highest scoring MS2 spectra was recorded and create a mirror plot comparing this MS2 spectra to the reference fragment spectra for caffeine.
par(mfrow = c(1, 2))
col_sample <- col_phenotype[sampleData(data)$phenotype]
#' Extract and plot EIC for the annotated feature
eic <- featureChromatograms(data, features = ms2_mtch_res$feature_id[1])
plot(eic, col = col_sample, peakCol = col_sample[chromPeaks(eic)[, "sample"]],
peakBg = paste0(col_sample[chromPeaks(eic)[, "sample"]], 20))
legend("topright", col = col_phenotype, legend = names(col_phenotype), lty = 1)
#' Identify the best matching query-target spectra pair
idx <- which.max(ms2_mtch_res$score)
#' Indicate the retention time of the MS2 spectrum in the EIC plot
abline(v = ms2_mtch_res$rtime[idx])
#' Get the index of the MS2 spectrum in the query object
query_idx <- which(query(ms2_mtch)$.original_query_index ==
ms2_mtch_res$.original_query_index[idx])
query_ms2 <- query(ms2_mtch)[query_idx]
#' Get the index of the MS2 spectrum in the target object
target_idx <- which(target(ms2_mtch)$spectrum_id ==
ms2_mtch_res$target_spectrum_id[idx])
target_ms2 <- target(ms2_mtch)[target_idx]
#' Create a mirror plot comparing the two best matching spectra
plotSpectraMirror(query_ms2, target_ms2)
legend("topleft",
legend = paste0("precursor m/z: ", format(precursorMz(query_ms2), 3)))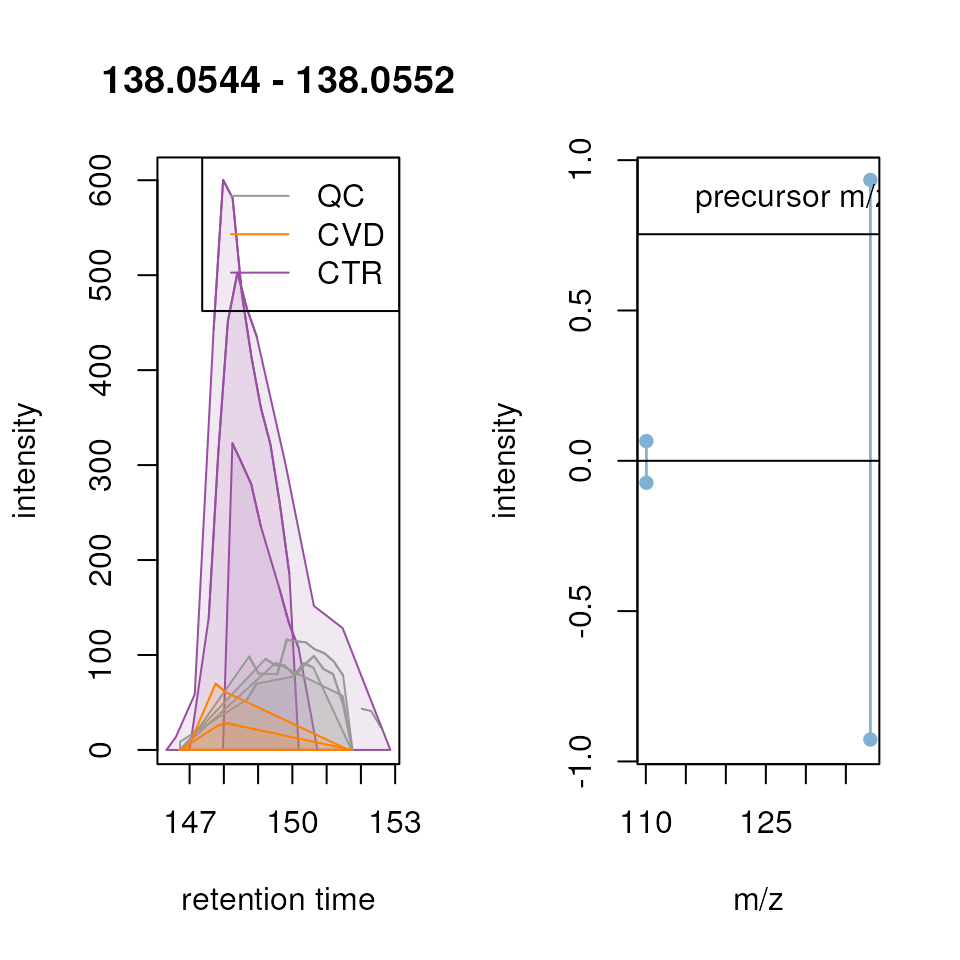
The plot above clearly shows the higher signal for this feature in CTR compared to CVD samples. The QC samples exhibit a lower but highly consistent signal, suggesting the absence of strong technical noise or biases in the raw data of this experiment. The vertical line indicates the retention time of the fragment spectrum with the best match against a reference spectrum. It has to be noted that, since the fragment spectra were measured in a separate LC-MS/MS experiment, this should only be considered as an indication of the approximate retention time in which ions were fragmented in the second experiment. The fragment spectrum for this feature, shown in the upper panel of the right plot above is highly similar to the reference spectrum for caffeine from MassBank (shown in the lower panel). In addition to their matching precursor m/z, the same two fragments (same m/z and intensity) are present in both spectra.
We can also extract additional metadata for the matching reference spectrum, such as the used collision energy, fragmentation mode, instrument type, instrument as well as the ion (adduct) that was fragmented.
spectraData(target_ms2, c("collisionEnergy_text", "fragmentation_mode",
"instrument_type", "instrument", "adduct")) |>
as.data.frame()## collisionEnergy_text fragmentation_mode instrument_type
## 1 55 (nominal) HCD LC-ESI-ITFT
## instrument adduct
## 1 LTQ Orbitrap XL Thermo Scientific [M+H]+External tools or alternative annotation approaches
The present workflow highlights how annotation could be performed
within R using packages from the Bioconductor project, but there are
also other excellent external softwares that could be used as an
alternative, such as SIRIUS (Dührkop et al.
2019), mummichog (Li et al. 2013)
or GNPS (Nothias et al. 2020) among
others. To use these, the data would need to be exported in a format
supported by these. For MS2 spectra, the data could easily be exported
in the required MGF file format using the
r Biocpkg("MsBackendMgf") Bioconductor package. Integration
of xcms into feature-based molecular networking with GNPS is
described in the GNPS
documentation.
In alternative, or in addition, evidence for the potential matching
chemical formula of a feature could be derived by evaluating the isotope
pattern of its full MS1 scan. This could provide information on the
isotope composition. Also for this, various functions such as the
isotopologues() from the or
r Biocpkg("MetaboCoreUtils") package or the functionality
of the envipat R package (Loos et al.
2015) could be used.
Summary
In this tutorial, we describe an end-to-end workflow for LC-MS-based untargeted metabolomics experiments, conducted entirely within R using packages from the Bioconductor project or base R functionality. While other excellent software exists to perform similar analyses, the power of an R-based workflow lies in its adaptability to individual data sets or research questions and its ability to build reproducible workflows and documentation.
Due to space restrictions we don’t provide a comprehensive listing of methodologies for the individual analysis steps. More advanced options or approaches would be available, e.g., for normalization of the data, which will however also be heavily dependent on the size and properties of the analyzed data set, as well as for annotation of the features.
As a result, we found in the present analysis a set of features with significant abundance differences between the compared groups. From these we could however only reliably annotate a single feature, that related to the lifestyle of individuals rather then to the pathological properties of the investigated disease. This low proportion of annotated signals is however not uncommon for untargeted metabolomics experiments and reflects the need for more comprehensive and reliable reference annotation libraries.
Session information
## R version 4.4.1 (2024-06-14)
## Platform: x86_64-pc-linux-gnu
## Running under: Ubuntu 22.04.5 LTS
##
## Matrix products: default
## BLAS: /usr/lib/x86_64-linux-gnu/openblas-pthread/libblas.so.3
## LAPACK: /usr/lib/x86_64-linux-gnu/openblas-pthread/libopenblasp-r0.3.20.so; LAPACK version 3.10.0
##
## locale:
## [1] LC_CTYPE=en_US.UTF-8 LC_NUMERIC=C
## [3] LC_TIME=en_US.UTF-8 LC_COLLATE=en_US.UTF-8
## [5] LC_MONETARY=en_US.UTF-8 LC_MESSAGES=en_US.UTF-8
## [7] LC_PAPER=en_US.UTF-8 LC_NAME=C
## [9] LC_ADDRESS=C LC_TELEPHONE=C
## [11] LC_MEASUREMENT=en_US.UTF-8 LC_IDENTIFICATION=C
##
## time zone: Etc/UTC
## tzcode source: system (glibc)
##
## attached base packages:
## [1] stats4 stats graphics grDevices utils datasets methods
## [8] base
##
## other attached packages:
## [1] MetaboAnnotation_1.9.1 CompoundDb_1.9.5
## [3] AnnotationFilter_1.29.0 AnnotationHub_3.13.3
## [5] BiocFileCache_2.13.0 dbplyr_2.5.0
## [7] gridExtra_2.3 ggfortify_0.4.17
## [9] ggplot2_3.5.1 vioplot_0.5.0
## [11] zoo_1.8-12 sm_2.2-6.0
## [13] pheatmap_1.0.12 RColorBrewer_1.1-3
## [15] pander_0.6.5 limma_3.61.11
## [17] MetaboCoreUtils_1.13.0 Spectra_1.15.10
## [19] xcms_4.3.3 BiocParallel_1.39.0
## [21] SummarizedExperiment_1.35.2 GenomicRanges_1.57.1
## [23] GenomeInfoDb_1.41.1 IRanges_2.39.2
## [25] S4Vectors_0.43.2 MatrixGenerics_1.17.0
## [27] matrixStats_1.4.1 MsBackendMetaboLights_0.99.0
## [29] MsIO_0.0.6 MsExperiment_1.7.0
## [31] ProtGenerics_1.37.1 readxl_1.4.3
## [33] Biobase_2.65.1 BiocGenerics_0.51.2
## [35] rmarkdown_2.28 knitr_1.48
## [37] BiocStyle_2.33.1
##
## loaded via a namespace (and not attached):
## [1] bitops_1.0-8 filelock_1.0.3
## [3] tibble_3.2.1 cellranger_1.1.0
## [5] preprocessCore_1.67.1 XML_3.99-0.17
## [7] lifecycle_1.0.4 doParallel_1.0.17
## [9] lattice_0.22-6 MASS_7.3-61
## [11] alabaster.base_1.5.9 MultiAssayExperiment_1.31.5
## [13] magrittr_2.0.3 sass_0.4.9
## [15] jquerylib_0.1.4 yaml_2.3.10
## [17] MsCoreUtils_1.17.2 DBI_1.2.3
## [19] abind_1.4-8 zlibbioc_1.51.1
## [21] purrr_1.0.2 RCurl_1.98-1.16
## [23] rappdirs_0.3.3 GenomeInfoDbData_1.2.12
## [25] MSnbase_2.31.1 pkgdown_2.1.1
## [27] ncdf4_1.23 codetools_0.2-20
## [29] DelayedArray_0.31.12 DT_0.33
## [31] xml2_1.3.6 tidyselect_1.2.1
## [33] farver_2.1.2 UCSC.utils_1.1.0
## [35] base64enc_0.1-3 jsonlite_1.8.9
## [37] iterators_1.0.14 systemfonts_1.1.0
## [39] foreach_1.5.2 tools_4.4.1
## [41] progress_1.2.3 ragg_1.3.3
## [43] Rcpp_1.0.13 glue_1.7.0
## [45] SparseArray_1.5.41 xfun_0.47
## [47] dplyr_1.1.4 withr_3.0.1
## [49] BiocManager_1.30.25 fastmap_1.2.0
## [51] rhdf5filters_1.17.0 fansi_1.0.6
## [53] digest_0.6.37 mime_0.12
## [55] R6_2.5.1 textshaping_0.4.0
## [57] colorspace_2.1-1 rsvg_2.6.1
## [59] RSQLite_2.3.7 utf8_1.2.4
## [61] tidyr_1.3.1 generics_0.1.3
## [63] prettyunits_1.2.0 PSMatch_1.9.0
## [65] httr_1.4.7 htmlwidgets_1.6.4
## [67] S4Arrays_1.5.9 pkgconfig_2.0.3
## [69] gtable_0.3.5 blob_1.2.4
## [71] impute_1.79.0 MassSpecWavelet_1.71.0
## [73] XVector_0.45.0 htmltools_0.5.8.1
## [75] bookdown_0.40 MALDIquant_1.22.3
## [77] clue_0.3-65 scales_1.3.0
## [79] png_0.1-8 reshape2_1.4.4
## [81] rjson_0.2.23 curl_5.2.3
## [83] cachem_1.1.0 rhdf5_2.49.0
## [85] stringr_1.5.1 BiocVersion_3.20.0
## [87] parallel_4.4.1 AnnotationDbi_1.67.0
## [89] mzID_1.43.0 vsn_3.73.0
## [91] desc_1.4.3 pillar_1.9.0
## [93] grid_4.4.1 alabaster.schemas_1.5.0
## [95] vctrs_0.6.5 MsFeatures_1.13.0
## [97] pcaMethods_1.97.0 cluster_2.1.6
## [99] evaluate_1.0.0 cli_3.6.3
## [101] compiler_4.4.1 rlang_1.1.4
## [103] crayon_1.5.3 labeling_0.4.3
## [105] QFeatures_1.15.3 ChemmineR_3.57.1
## [107] affy_1.83.1 plyr_1.8.9
## [109] fs_1.6.4 stringi_1.8.4
## [111] munsell_0.5.1 Biostrings_2.73.2
## [113] lazyeval_0.2.2 Matrix_1.7-0
## [115] hms_1.1.3 bit64_4.5.2
## [117] Rhdf5lib_1.27.0 KEGGREST_1.45.1
## [119] statmod_1.5.0 highr_0.11
## [121] mzR_2.39.0 igraph_2.0.3
## [123] memoise_2.0.1 affyio_1.75.1
## [125] bslib_0.8.0 bit_4.5.0Appendix
Additional informations
#possible extra info:
# -